


1. Introduction
The ability of organisms to defend themselves against external pathogens and to repair tissue injury brought on by infection is vital for them to survive. Inflammatory processes can contribute to the regulation of the origin, development, and outcomes of autoimmune and autoinflammatory diseases [1]. Moreover, inflammatory responses can damage host tissues and result in organ failure in a wide range of disorders. These inflammatory diseases are characterized by inflammation, which has been identified as the leading cause of death worldwide [2].
Innate immune responses serve as the initial line of defense for the host against pathogens. These responses can help detect and eliminate infected cells as well as coordinate and activate the development of adaptive immunity [3,4]. Neutrophils play a crucial role as effector cells in the innate immune system. Neutrophils are considered a type of polymorphonuclear (PMN) leukocyte. They are emerging as highly specialized cells that are capable of carrying out a number of immune defense-related functions [5]. Neutrophils constitute the most prevalent leukocytes in circulation and possess a short lifespan. If microbial infections are detected, these cells immediately act to trap and eliminate invasive pathogens [6]. In addition, neutrophils interact in complicated, bidirectional ways with certain other immune cells once they reach inflamed tissues, affecting both innate and adaptive immune responses [7].
Furthermore, neutrophils are naturally the first to respond to acute inflammation and aid in its resolution. Neutrophils have additionally been proven to be important in chronic inflammation throughout the past few decades. Neutrophils are persistently recruited towards the site of chronic inflammation. They help drive inflammatory processes by releasing inflammatory factors and several cytokines that can regulate inflammation and activate other types of immune cells [8,9]. Numerous pathological conditions such as cancer, neurological, metabolic, and autoimmune disorders are caused by neutrophil-mediated inflammation [10].
Moreover, neutrophils were previously considered a homogeneous population with conserved phenotypes and distinct roles. Current studies have shown that heterogeneous populations of neutrophils have diverse functional phenotypes, especially in pathological conditions of inflammation [11]. Because neutrophils serve as primary effector cells, the severity and type of inflammatory response after severe injury may be determined by the expression pattern of neutrophil receptors. Neutrophil heterogeneity might serve as a helpful risk assessment tool [12,13].
Many different approaches targeting neutrophils, including strategies that use a number of different agents to improve, hinder, or restore neutrophil activity, have emerged. However, there are still challenges and controversies that remain in the field of neutrophil research. This review emphasizes the pathogenic role of neutrophils and their derived molecules, elucidating their relation to various inflammatory diseases. It also outlines novel insights into phenotypical and functional heterogeneity of neutrophils during inflammatory diseases. In addition, some effective potential therapeutic attempts that specifically target neutrophils are summarized.
2. Neutrophil Activation
Neutrophil activation is often a multi-step process that is advantageous for killing pathogens. However, neutrophil activation pathways can also potentially cause tissue damage in autoimmune and inflammatory diseases. Some stimulants, including pathogen chemoattractants (fMLP) and bacterial lipopolysaccharide (LPS), are important for activating neutrophils [14,15]. These stimulants can adhere to neutrophils and activate their receptors to enhance responsiveness of those cells to subsequent stimuli. There are several different kinds of receptors that are expressed on neutrophils, including G-protein-coupled receptors (GPCRs), Fc-receptors, adhesion molecules/receptors such as integrins and selectins/selectin ligands, different cytokine receptors, and innate immune receptors such as C-type lectins and toll-like receptors (TLRs). Activation of such receptors can cause additional reactions such as chemotactic migration or the release of chemokines and cytokines as well as multiple cellular activation and eradication processes such as phagocytosis, generation of reactive oxygen species (ROS), exocytosis of intracellular granules, and the releasing of neutrophil extracellular traps (NETs).
GPCRs like formyl peptide receptors (FPR1, FPR2) have been recognized as bacterial products and mitochondrial peptides via the MAPK/ERK pathway [16], leukotriene B4 (LTB4), platelet-activating factor receptors (PAFRs), complement receptors (C3aR, C5aR1), CXC chemokine receptors (CXCR1, CXCR2, CXCR3, CXCR4), and, to a smaller extent, CC chemokine receptors (CCR1, CCR2, CCR3, CCR5, ACKRs, CCRL2). These GPCRs are capable of enhancing neutrophil responses to further activation. Furthermore, the chemotactic migratory activity of neutrophils is substantially triggered by these receptors [17,18,19]. Several Fc-receptors that are expressed by neutrophils (Fcγ, Fcε, Fcα) have a role in the identification of immunoglobulin (Ig)-opsonized pathogens. They are also involved in immune-mediated inflammatory conditions. Low-affinity Fcγ-receptors are among the most significant Fc-receptors of neutrophils [20,21]. Additionally, neutrophil activation often happens when they bind to extracellular matrix proteins or other cells through integrin or selectin adhesion receptors. Neutrophils can adhere firmly when integrins interact with their specific ligands, such as intercellular adhesion molecules (ICAMs) on endothelial cells. Several kinases, including Src-family kinases, phosphoinositide 3-kinase (PI3K), tyrosine kinase (SYK), and p38 mitogen-activated protein kinase (MAPK), are activated by β2 integrin ligation and selectin–selectin ligand interaction [22,23,24,25]. The activation of these kinases is important for neutrophil functions. Numerous cytokine receptors are also expressed by neutrophils, including tumor necrosis factor (TNF) receptors, granulocyte-colony stimulating factor (G-CSF), and granulocyte macrophage-colony stimulating factor (GM-CSF) receptors, as well as different interleukins (ILs) and interferons (IFN) receptors [22]. These receptors control a variety of neutrophil functions through intercellular communication.
Numerous innate immune receptors, also known as pattern recognition receptors (PRRs), are expressed on neutrophils. They have a direct role in the identification of microbes and tissue injury. According to the homology of protein domains, PRRs can be divided into the following categories: TLRs, retinoic acid-inducible gene-I (RIG-I)-like receptors (RLRs), nucleotide-oligomerization domain (NOD)-like receptors (NLRs) and C-type lectin receptors (CLRs) [22,26,27]. In neutrophils, activation of these receptors is possible through a variety of signal transduction mechanisms. Neutrophil PRRs are important regulators of host immunological responses.
Consequently, these neutrophils’ stimulated receptors can detect microbes as well as an inflammatory environment. Understanding these neutrophil receptors and related signaling pathways that regulate neutrophil function is necessary for the development of therapies that can prevent host tissues from being harmed by neutrophils.
3. Structure and Functions of Neutrophils
Neutrophils are composed of different granule types: (1) primary granules, commonly referred to as azurophilic granules; (2) secondary granules, also called specific granules; (3) tertiary granules, also known as gelatinase granules; and (4) secretory granules. Primary granules contain most mediators, including elastase, MPO, proteinase 3, Cat-G, azurocidin, and defensins. Secondary granules are the storage site of lactoferrin, cathelicidin, neutrophil gelatinase-associated lipocalin (NGAL), and collagenase. Tertiary granules comprise matrix metalloproteases (MMPs). Secretory granules contain plasma proteins and cationic antimicrobial protein 37 (CAM37). Activated neutrophils can also release other effector molecules such as reactive oxygen species (ROS), LTB4, calprotectin, peptidyl arginine deiminase (PAD), NETs, and various types of cytokines and chemokines, as shown in Figure 1.
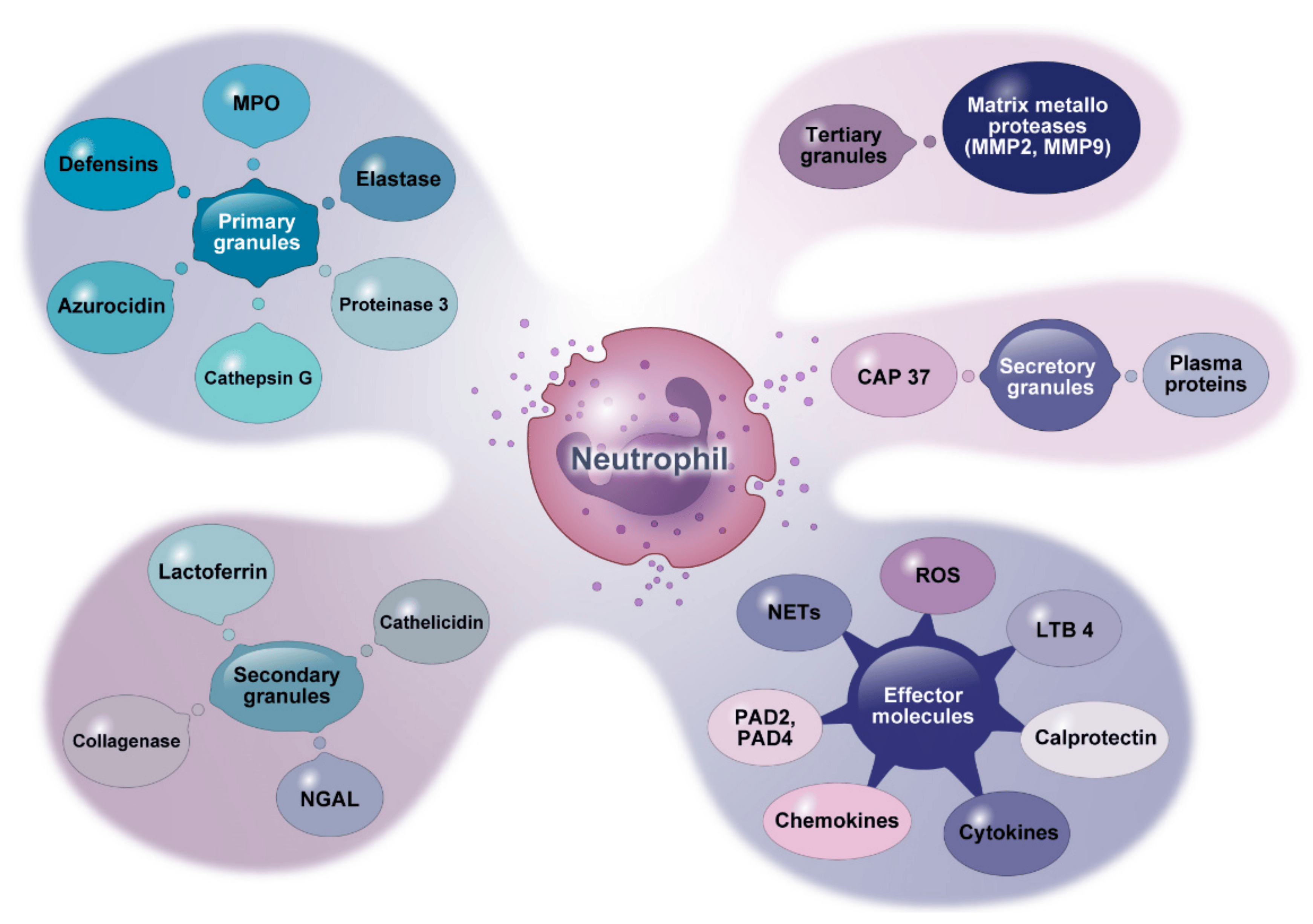
Figure 1. Granule contents and other molecules secreted by neutrophils. This figure shows effective mediators derived from neutrophil granules and other effector molecules they release. MPO: myeloperoxidase. NGAL: neutrophil gelatinase-associated lipocalin. CAP: cationic antimicrobial protein. LTB4: leukotriene B4. PAD: peptidyl arginine deiminases.
Neutrophils are multifaceted cells with a wide range of distinct functions. Appropriate neutrophil recruitment is required for modulation and resolution of inflammation, tissue repair, wound healing, elimination of microorganisms, and restoration of homeostasis, as depicted in Figure 2. These various roles that neutrophils play will be discussed below.
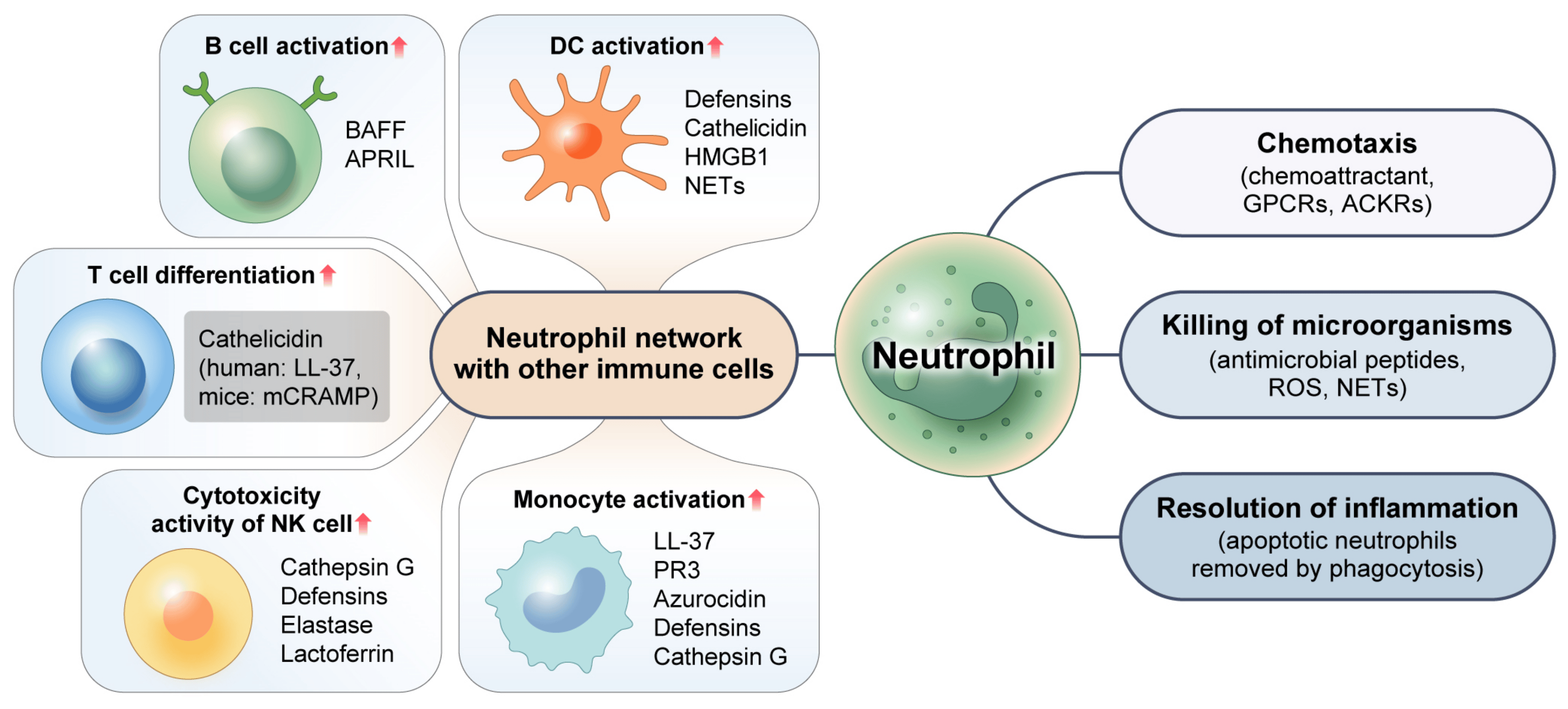
Figure 2. Multifaceted function of neutrophils. Neutrophils eradicate intracellular pathogens through their chemotaxis activity and the release of antimicrobial peptides and ROS. By efficiently engulfing and removing apoptotic neutrophils via the process of phagocytosis, neutrophils can facilitate the resolution of inflammation. Neutrophils can also promote immune responses via complex interactions with other immune cells and use various mechanisms to control activities of other immune cells. GPCRs: G protein-coupled receptors. ACKRs: atypical chemokine receptors. ROS: reactive oxygen species. NETs: neutrophil extracellular traps. HMGB1: high-mobility group box 1. BAFF: B-cell activating factor. APRIL: a proliferation-inducing ligand. PR3: proteinase 3.
3.1. Chemotaxis
Neutrophils can identify inflammatory signs and move in the direction of infected sites [28]. For this efficient response, they can recognize extracellular chemical gradients and migrate toward higher concentrations through a process known as chemotaxis. Chemoattractants are a set of molecular guidance cues with diverse chemical compositions. They are responsible for orchestrating this targeted neutrophil recruitment. In humans, these chemoattractant molecules can be classified into four molecular families: formyl peptides, chemokines, anaphylatoxins, and chemotactic lipids [29]. Neutrophil chemoattractants work by interacting with heptahelical GPCRs expressed on cell surfaces. The Rho family of GTPases play a major role in controlling chemotactic response. Moreover, emerging data indicate that the atypical chemoattractant receptor (ACKR), another receptor that does not bind to G proteins, might also play a significant role in regulating the migratory ability and functional responses of neutrophils. The expression of chemoattractant receptors is reliant on the degree of neutrophil maturation state and activation, with a crucial modulatory function for an inflammatory condition [17].
3.2. Killing of Microbes
Neutrophils can phagocytose microorganisms at sites of infection. Throughout this process, both primary and secondary granules combine well with phagosomes and produce proteins and antimicrobial peptides such as myeloperoxidase (MPO), neutrophil elastase (NE), cathepsin G (Cat-G), cathelicidin, alpha-defensins, and many others. During the same periods, ROS are generated via nicotinamide adenine dinucleotide-phosphate (NADPH). NETs are produced by activated neutrophils in response to specific stimuli. NETs resemble a net-like network made up of cell-free DNA, neutrophil granule proteins, and histones. NETs are recognized as a pathogen control strategy identified only a few years ago [30]. All of these microbicidal molecules are secreted by neutrophils to create a highly dangerous environment that seems to be necessary for effective microbial killing and destruction. Several of these microbicidal molecules can aid in the development of infection by being cytotoxic to host tissues [31,32]. Thus, it is expected that the host uses a variety of mechanisms to restrict or stop them from harming host cells and causing undesired inflammation.
3.3. Resolution of Inflammation
Neutrophils have a short lifespan. They are limited by apoptosis. Apoptotic neutrophils are functionally inactive due to a programmed shutdown and disabling of their signaling pathways. However, they can also retain cell-surface receptors that allow them to be recognized and phagocytosed by macrophages and other phagocytic cells. By removing apoptotic neutrophils from inflammatory areas, it is possible to avoid tissue damage that might otherwise occur due to the discharge of cytotoxic compounds into surrounding tissues that might have died due to necrosis. The efficient death of neutrophils and the secure evacuation of apoptotic neutrophils through phagocytic cells are crucial for the reduction in inflammation [9,33]. Under inflammatory conditions, disruption of neutrophil apoptosis can result in prolonged survival of neutrophils in damaged tissue and prolong the secretion of neutrophil-derived immunomodulatory cytokines, cytotoxic chemicals, and chemokines, which may lead to prolonged inflammation [34,35]. Failure in the removal of apoptotic neutrophils could also result in the generation of autoantibodies, because these cells express autoantigens upon their surface [36].
3.4. Neutrophil Network with Other Immune Cells
Neutrophils not only participate in the eradication of microbes, but also promote immune reactions to intracellular pathogens through intricate interactions with other immune cells. For instance, neutrophils can release chemokines (such as CCL3, CCL4, CCL5, and CCL20) and alarmins, including α-defensins, cathelicidins, and high-mobility group box-1 (HMGB1) proteins that are chemotactic for dendritic cells (DCs). These chemokines and alarmins are required for efficient DC recruitment to infected sites. NETs can promote plasmacytoid (p) DCs to release inflammatory cytokines [37]. Similarly, interactions between macrophages and neutrophils play a crucial role in both the beginning and resolving stages of an inflammatory reaction. Tissue-resident macrophages can secrete some chemoattractants like CXCL1, CXCL2, CCL2, and IL-1α that are required for the migration of activated neutrophils toward the inflammatory site. This process can increase the lifespan of neutrophils by secreting G-CSF, GM-CSF, and TNF-α [38]. When neutrophils arrive at the site of inflammation, they can activate the immune system by recruiting monocytes and releasing proteins such as LL-37, proteinase 3 (PR3), azurocidin, defensins, and Cat-G [39,40]. Defensins, azurocidin, and other antimicrobial peptides can boost antimicrobial activities of macrophages by enhancing their capacity to phagocytose and generate cytokines (TNF-α and IFN-γ) [41,42].
Cytokines BAFF (B-cell activating factor) and APRIL (a proliferation-inducing ligand) are generated by neutrophils in great quantities. These cytokines are essential for B-cell survival, development, and differentiation. They are elevated in inflammatory diseases [43,44]. Furthermore, the function of several T-cell subsets can be positively or negatively modulated by neutrophils. In both humans and animals, activated neutrophils encourage T-cell activation, multiplication, and differentiation into effector CD8+ T-cells, and T-helper cell subsets (Th1, Th17) could promote adaptive responses at the inflammatory site [45,46,47]. According to previous studies, granule peptides of neutrophils such as cathelicidin (mCRAMP in mice, LL-37 in humans) can exert immunomodulatory actions on T cells and modulate Th1 and Th17 differentiation [48]. At the site of inflammation, neutrophils can also stimulate natural killer (NK) cells. Cat-G, defensins, elastase, and lactoferrin (LTF) are implicated in the augmentation of cytotoxic activity of human NK cells. The survival rate of human neutrophils can be improved by NK-derived substances including GM-CSF and IFN-γ [49]. These findings suggest that human neutrophils have a variety of mechanisms through which they can control the activity of NK cells.
Crosstalk between neutrophils and other immune cells as well as significant chemical signals can affect the development and remission of inflammation. However, further research on associations of neutrophils with other immune cells in relation to inflammatory diseases is required.
4. Neutrophils in Infection
Neutrophils are essential mediators that can serve in the initial defense against invasive pathogens like bacteria and viruses. Next, we will explain the effector role of neutrophils in bacterial and viral infections.
4.1. Bacterial Infection
Neutrophils are required as a key component of the innate response to bacterial infection. Upon bacterial infection, neutrophils will exit the bloodstream and move to the inflammatory area to fight against bacterial infections. When an infection is caused by Listeria monocytogenes, a Gram-positive intracellular pathogen, neutrophils can move from the bone marrow to the infectious site. They employ unique bacterial-sensing mechanisms at this site that can result in phagocytosis and generation of bactericidal substances [50]. Liu et al. [51] have investigated formyl peptide receptors related to chemoattractant GPCRs which are crucial for the quick migration of neutrophils in Listeria-infected livers of mice for successful clearance of infectious microbes. LTB4, another chemoattractant for neutrophils, is essential for neutrophil colonization. A recent publication has shown that preincubation of human neutrophils with the Gram-negative intracellular bacteria Salmonella typhimurium can promote neutrophil colonization [52]. This bacterium has been used to stimulate LTB4 production caused by a bacterial chemoattractant fMLP, which is important for the eradication of pathogens instantaneously. However, certain bacterial infections also generate molecules that can inhibit neutrophil recruitment. Extracellular bacteria like Streptococcus pyogenes (S. pyogenes) and Streptococcus pneumoniae (S. pneumoniae) use a significantly different strategy to prevent neutrophil recruitment. For example, streptolysin is an effective cytolytic toxin produced by S. pyogenes. This streptolysin is essential for inhibiting neutrophil recruitment in the initial stages of S. pyogenes infection, as observed in studies using zebrafish [53]. Similarly, zinc metalloproteinase produced by S. pneumoniae can cleave P-selectin glycoprotein 1 (PSGL-1), preventing neutrophil extravasation in its first stages [54]. Moreover, Staphylococcus aureus is an intracellular bacterium with chemotaxis inhibitory protein that has been employed to prevent neutrophil activation [53,55,56,57].
Additionally, phagocytosis, a mechanism that occurs in neutrophils, allows them to eliminate bacterial infections. There are multiple examples of bacteria that can secrete different compounds to either increase or decrease the phagocytic activity of neutrophils. For instance, Neisseria gonorrhoeae can alter mitochondrial depolarization and caspase activation to regulate phagocytosis in human neutrophils [58]. In contrast, S. pneumoniae capsules can reduce bacterial opsonization and inhibit effective recognition by complement receptors, Fcγ receptors, and nonopsonic receptors. Due to this inhibition, neutrophils are unable to phagocytose the bacterium, which allows S. pneumoniae to cause diseases [59]. Other bacteria including Mycobacterium tuberculosis, Neisseria meningitidis, Haemophilus influenzae, Pseudomonas aeruginosa, and Escherichia coli can also inhibit the phagocytic activity of neutrophils through different mechanisms [57,60,61,62].
Moreover, neutrophil apoptosis is a pro-resolution process that can reduce the severity of tissue damage and inflammation. However, accelerating or delaying neutrophil apoptosis might have several negative consequences. Some bacterial pathogens such as Pseudomonas aeruginosa can release a pigment known as pyocyanin and exotoxin A that can cause apoptosis of neutrophils [63]. Other examples of bacteria that can persuade apoptosis of neutrophils following phagocytosis include Salmonella typhimurium, Escherichia coli, and Staphylococcus aureus [64,65,66,67]. Numerous bacteria have been proven to cause neutrophil apoptosis, although fewer bacteria have been found to be able to prevent this death process. For example, Chlamydia psittaci, Francisella tularensis, and Anaplasma phagocytophilum are other intracellular bacterial pathogens that can delay neutrophil apoptosis via signaling pathways and anti-apoptotic proteins.
NETs are produced by neutrophils through a process known as NETosis, which traps a variety of bacteria. The immunological response to bacterial infections is highly dependent on NETs. NETs can inhibit the growth of bacteria including Shigella flexneri, Escherichia coli, Pseudomonas aeruginosa, Salmonella typhimurium, Shigella sonnei, Klebsiella pneumoniae, Salmonella enteritidis, Staphylococcus albus, Pseudomonas aeruginosa, Propionibacterium, and Staphylococcus aureus and kill them [68,69].
Overall, the communication between neutrophils and bacteria is dynamic and complex. These are only a few examples. There are undoubtedly many more bacteria that can affect neutrophils. The role of neutrophils during various other bacterial infections is still largely unexplored. Understanding the role of neutrophils in the defense against bacterial infection can be extremely helpful in the development of new therapies for bacterial infection.
4.2. Viral Infection
Neutrophils have a variety of functions in severe viral infections. They can limit viral replication and transmission by phagocytosis, respiratory burst, degranulation, cytokine production, antimicrobial peptides, formation of NETs, and activation of the adaptive response. However, excessive activation of neutrophils can harm the tissue with negative effects. Many viruses such as herpes simplex virus (HSV) [70], respiratory syncytial virus (RSV) [71], influenza A virus (IAV) [72], and human immunodeficiency virus (HIV) [73] can activate neutrophils via PRRs to release proinflammatory cytokines, chemokines, ROS, and granular enzymes. Furthermore, Cloke et al. [74] have revealed that certain neutrophil phenotypes like low-density granulocytes (LDGs) are associated with certain viral infections. In HIV-positive patients, LDGs are presumably activated neutrophils that are primed to degranulate. Contrarily, hepatitis C virus (HCV) can decrease neutrophil phagocytosis in both cirrhotic and non-cirrhotic individuals, indicating that neutrophil dysfunction is associated with HCV replication [75]. Research on mice infected with IAV has revealed that blocking C5a can reduce neutrophil recruitment to lungs and tissue damage [76].
It has been demonstrated that many viruses can directly or indirectly promote or prevent neutrophil apoptosis. For instance, the influenza virus can accelerate neutrophil apoptosis. Studies have demonstrated that exposure to the influenza virus might cause neutrophils to upregulate pro-apoptotic factors like Fas and TNF-related apoptosis-inducing ligands (TRAIL). As a result, caspase-8 and caspase-3 are activated, starting the apoptotic cascade to help eliminate infected cells [77]. Several viruses such as HCV, HIV, and Simian immunodeficiency virus (SIV) have been found to increase neutrophil apoptosis. These viruses can cause neutrophil apoptosis by a variety of complex mechanisms/pathways, including the secretion of ROS and cytokines [78,79,80]. When these viruses cause enhanced apoptosis of neutrophils, they can induce neutropenia, potentially weakening the immune response to infection [81,82]. In contrast, delaying neutrophil apoptosis has the potential to worsen tissue damage and accelerate viral clearance. Therefore, the host may benefit from suppression of neutrophil apoptosis. For instance, it has been revealed that human cytomegalovirus (HCMV) can inhibit neutrophil apoptosis and cause production of a highly bioactive secretome (TNF-α, IL-6, IL-8, MIP-1α, IL-13, and IL-10) that can promote neutrophil survival and trigger the chemotaxis of monocytes and their differentiation into permissive, anti-inflammatory phenotypes [83]. In addition, one research paper has demonstrated that RSV can also extend the longevity of human neutrophils by preventing or delaying apoptosis [84].
Likewise, NETs can trap and eliminate viral infections. Certain viruses, including influenza A, HIV-1, and RSV, can cause the development of NETs. By generating ROS species and activating TLRs 4, 7, or 8, these viruses can trigger NETosis, a process by which NETs can trap and destroy viruses [85,86,87,88]. Nevertheless, acute viral infections such as those brought by dengue virus (DV) and coronavirus-2 are known to generate dysregulated NETs. Dysregulated NET formation has been demonstrated to be a measure of disease severity. It plays a part in the development of infection [89].
In summary, viruses can affect neutrophil activities in a variety of ways, which can affect immune responses and aid in the emergence of viral infections. Awareness of complex interactions among neutrophils and viruses can offer useful insights for the development of efficient therapies against viral diseases.
5. Role of Neutrophils in Inflammatory Diseases
Inflammatory diseases are defined by chronic or persistent inflammatory responses that can cause tissue injury and malfunction. Neutrophils, their derived molecules, and neutrophil heterogeneity all have an effective role in the emergence and perpetuation of inflammatory diseases, including multiple sclerosis (MS), inflammatory bowel disease (IBD), Behçet’s disease (BD), atopic dermatitis (AD), rheumatoid arthritis (RA), and systemic lupus erythematosus (SLE), as shown in Figure 3 [32,90]. Our expanding knowledge of neutrophil function in different inflammatory diseases might have a considerable impact on the development of targeted treatments for inflammatory diseases.
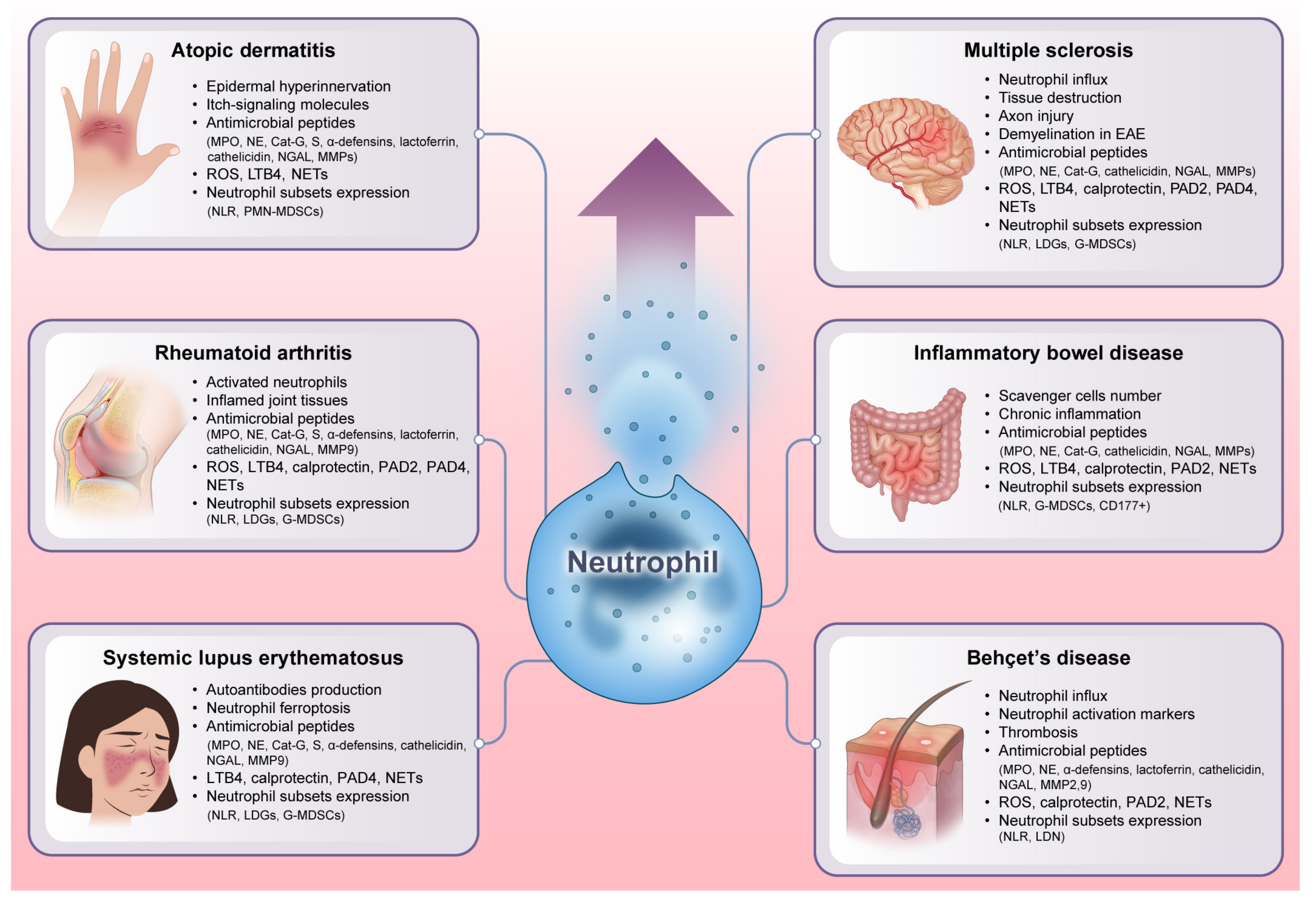
Figure 3. Unraveling the explosive role of neutrophils in inflammatory diseases. Neutrophils exert a profound impact on the development and maintenance of inflammation by secreting antimicrobial peptides and presenting diverse heterogeneity. As immunological orchestrators, neutrophils can also release many other powerful chemicals such as cytokines, chemokines, ROS, and NETs that can intensify the inflammatory cascade. Additionally, the variability of neutrophil subsets, each of which has unique functional characteristics and phenotypes, contributes to the variety of symptoms and severity seen in different inflammatory diseases. All of these factors emphasize that neutrophils can act as vital players in disease pathogenesis. EAE: experimental autoimmune encephalomyelitis. NLR: neutrophil-lymphocyte ratio. LDGs: low-density granulocytes. MDSCs: myeloid-derived suppressor cells. CD: a cluster of differentiation.
5.1. Multiple Sclerosis
MS is an immune-mediated, demyelinating, chronic inflammatory, and neurodegenerative disease of the central nervous system (CNS). It has an unknown etiology. Although the majority of inflammatory cells related to MS are macrophages and T lymphocytes that aggregate inside perivascular regions and brain parenchyma, evidence shows that neutrophils also have a negative impact on the development of MS [91,92]. Recent research has shown that in the early phase of the disease, levels of neutrophils and other leukocytes are much higher in MS patients [93]. During MS, neutrophils can infiltrate into the CNS and lead to tissue destruction and inflammation, which are hallmarks of the disease [94]. Moreover, in the latest genome-wide association study (GWAS) of MS, neutrophil cytosolic factor-4 (NCF-4) gene, encoding one of the subunits of the nicotinamide-adenine dinucleotide phosphate (NADP) complex in neutrophils, was discovered as a genetic factor susceptible to MS [95]. Moreover, neutrophils can promote immune-mediated demyelination in experimental autoimmune encephalomyelitis (EAE), a murine MS model [96]. Collectively, neutrophils exhibit a wide range of effector actions that facilitate the disease pathogenesis.
5.2. Inflammatory Bowel Disease
The most common diseases among BD are Crohn’s disease (CD) and ulcerative colitis (UC). They are mainly characterized by severe inflammation of the gastrointestinal (GI) tract. The actual origins of IBD are unknown. Studies have identified pathogenic immune cell networks and abnormal immune cell trafficking as crucial drivers of tissue damage and mucosal inflammation in IBD [125]. Among various immune cells, the migrating activity of neutrophils toward the colon mucosa is a specific feature of IBD. Neutrophils are thought to perform dual functions in IBD. Firstly, they move toward the intestinal lining to assist in the defense against dangerous bacteria and other infectious agents. Secondly, neutrophils are constantly accumulating or becoming active inside the intestinal mucosa. They can exceed the number of scavenger cells. Prolonged and excessive activation of neutrophils might result in chronic inflammatory processes in IBD [126,127]. However, the exact role of neutrophils in IBD is unclear yet and appears to vary depending on the experimental conditions.
5.3. Behçet’s Disease
BD is known as an inflammatory disease with numerous manifestations. It can be recognized by the involvement of the vascular, articular, neurologic, and gastrointestinal systems as well as ocular, skin, genital, and oral ulcers [165]. Although the exact etiology of BD is unidentified, immunological irregularities play significant roles in the pathogenesis. It is well known that the pathogenesis of BD is associated with increased neutrophil influx and disturbed self-tolerance [166,167]. Neutrophils usually participate in perivascular infiltration in lesions and show significant intrinsic activity in BD patients. Chemotaxis and phagocytosis can both be increased by hyperactive neutrophils [168,169]. These heightened neutrophil functions contribute to tissue damage and immunological alterations observed in the disease. Patients with active BD have higher levels of neutrophil activation markers including CD64 [170]. Furthermore, neutrophil inflammation is a major mediator of thrombosis in BD. There are also reports of circulating neutrophil impairments in BD due to activation-induced cell death (AICD). A histological study of BD lesions has revealed arterial and venous infiltrates of neutrophils, indicating that neutrophils are specifically implicated in BD lesions [171]. It has been suggested that BD can be categorized as neutrophilic vasculitis [172,173]. Overall, neutrophils are mediators of inflammation that occur in BD patients.
5.4. Atopic Dermatitis
AD is a chronic inflammatory skin disease that causes extremely itchy redness and inflamed skin. The most prevalent feature of atopic diseases is immune globulin E (IgE)-mediated allergic reactions related to environmental allergens. AD is considered a heterogeneous disorder with a spectrum of morphology, dispersion, and disease progression. The pathophysiology is complicated, and numerous cell types are involved, including immune cells, skin cells, and neuronal cells that monitor and control immune responses [199,200]. Neutrophils are the initial immune cells that infiltrate AD skin. Moreover, neutrophils can regulate early skin hyperinnervation and upregulate the expression of activity-induced genes and itch-signaling molecules in nerve cells [201,202]. These findings imply that neutrophils might influence AD onset and progression via a variety of mechanisms.
5.5. Rheumatoid Arthritis
Rheumatoid arthritis (RA) is an autoimmune, chronic, and heterogeneous disease. It can be defined by an increasing symmetric joint inflammation that causes bone erosion, cartilage damage, and impairment. It has recently been clear that RA develops from epigenetic, genetic, and environmental triggers, although immunological variables must also play a significant role [230,231]. The pathophysiology of RA is influenced by dysregulation of neutrophil activity. Neutrophils, the most prevalent leukocytes in affected joints, are crucial for the development and persistence of RA [232,233]. Both synovial fluid (SF) and synovial tissue (ST) from RA joints contain a high concentration of activated neutrophils [234]. Activated neutrophils play a role in inflammation and damage to host tissues through degranulation, which occurs either into the SF or directly onto the joint surface. Neutrophils can migrate to the joint. The migration of neutrophils to the joint is a defining feature of inflammatory arthritides5, notably RA. Neutrophils could also serve as a reservoir for autoantigens that instigate the autoimmune processes that underlie this pathogenic condition. A comprehensive grasp of the intricate involvement of neutrophils in RA is imperative for addressing this persistent autoimmune condition.
5.6. Systemic Lupus Erythematosus
Systemic lupus erythematosus (SLE) is a chronic, complex, prototypical autoimmune, and worldwide disease. SLE is influenced by environmental, endocrine, immune, and genetic predispositions. Moreover, the body’s immune system overproduces autoantibodies, which results in widespread tissue damage and inflammation. Increased autoantibodies and accumulation of immune complexes are major hallmarks of SLE patients [269,270,271]. Up to this point, the majority of pathology research has concentrated on abnormalities of adaptive immune responses. However, the pathophysiology of SLE has currently received significant interest in innate immune responses, which operate earlier, before adaptive immune responses. Particularly, neutrophils exhibit multiple aspects of dysregulation. Neutrophils exhibit epigenetic alterations and genomic modifications. Furthermore, neutrophils play a major role in the development of SLE by encouraging exposure to self-antigens and generation of autoantibodies. Recently, it has been found that neutrophil ferroptosis has a significant pathogenic impact on SLE. Moreover, interferonopathies, which are defined by excessive type 1 IFN production due to genetic mutation, have a crucial role in SLE. This dysregulated type 1 IFN production promotes the generation of autoantibodies, which is an important aspect of SLE [272]. Dysregulated innate immunity is sufficient to upset the balance of immunological tolerance [273,274,275]. Overall, neutrophils play a complicated role in SLE.
6. Therapeutic Interventions Targeting Neutrophil-Derived Molecules in Inflammatory Diseases
According to findings outlined in previous sections, neutrophils have a vital role in inflammatory conditions. Uncontrolled or inappropriate activity of neutrophils could be a factor in tissue damage in inflammatory conditions and other diseases. Moreover, excessive release of neutrophil-derived molecules and neutrophil heterogeneity can promote inflammatory responses in neutrophil-mediated diseases. In such circumstances, therapeutic approaches to target neutrophils would be able to suppress neutrophil activity by inhibiting excessive secretion of neutrophil-derived molecules. As listed in , specific targets and inhibitors can be used to modulate neutrophil function in disease models such as MS, AD, IBD, BD, RA, and SLE. Nevertheless, future studies are still needed to reduce treatment side effects and improve function.
7. Conclusions
Neutrophils are considered vital cells of our innate immunity with a main function in host defense. Besides being a significant player in innate immunity, a growing body of research suggests that neutrophils have a variety of functions in many infectious and inflammatory diseases. The main way neutrophils aggravate disease is by producing neutrophil-derived molecules and NETs. Neutrophil-derived molecules, ROS, and NETs have a double-edged effect on the severity of many inflammatory diseases. Furthermore, neutrophils demonstrate a wide variety of phenotypes. Recent evidence reveals the presence of specific neutrophil heterogeneity in inflammatory diseases. Nonetheless, their properties, prevalence, and pathological potential require further investigation. Although there have been immense advances in our awareness of the functions of neutrophils in diseases, there is still more to learn regarding the mechanisms behind the migration of neutrophil-derived molecules to different tissues and the function of neutrophil heterogeneity in pathological conditions.
Moreover, several researchers have concentrated their attempts on targeting neutrophil activity as a potential treatment. Many inhibitors and compounds have been recognized as therapeutics by targeting neutrophils over the years. Despite that, more studies should focus on limiting negative effects of inhibitors and substances used for preventing massive release of neutrophil-derived molecules and NET formation. Furthermore, reducing or blocking neutrophil heterogeneity needs additional knowledge of their origin and function in inflammatory diseases and immunological defense. Altogether, understanding neutrophil activity, neutrophil-mediated pathogenesis, neutrophil-derived molecules, and neutrophil heterogeneity is imperative. These considerations, by particularly targeting pathogenic neutrophils without adversely affecting immunity, may therefore be crucial in the development of novel therapies for inflammatory diseases.
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.A.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1736–1788. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, X.; Saredy, J.; Yuan, Z.; Yang, X.; Wang, H. Innate-adaptive immunity interplay and redox regulation in immune response. Redox Biol. 2020, 37, 101759. [Google Scholar] [CrossRef] [PubMed]
- Riaz, B.; Islam, S.M.S.; Ryu, H.M.; Sohn, S. CD83 Regulates the Immune Responses in Inflammatory Disorders. Int. J. Mol. Sci. 2023, 24, 2831. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The multifaceted functions of neutrophils. Annu. Rev. Pathol. 2014, 9, 181–218. [Google Scholar] [CrossRef] [PubMed]
- Leliefeld, P.H.; Koenderman, L.; Pillay, J. How Neutrophils Shape Adaptive Immune Responses. Front. Immunol. 2015, 6, 471. [Google Scholar] [CrossRef]
- Herrero-Cervera, A.; Soehnlein, O.; Kenne, E. Neutrophils in chronic inflammatory diseases. Cell. Mol. Immunol. 2022, 19, 177–191. [Google Scholar] [CrossRef]
- Greenlee-Wacker, M.C. Clearance of apoptotic neutrophils and resolution of inflammation. Immunol. Rev. 2016, 273, 357–370. [Google Scholar] [CrossRef]
- Hedrick, C.C.; Malanchi, I. Neutrophils in cancer: Heterogeneous and multifaceted. Nat. Rev. Immunol. 2022, 22, 173–187. [Google Scholar] [CrossRef]
- Hellebrekers, P.; Vrisekoop, N.; Koenderman, L. Neutrophil phenotypes in health and disease. Eur. J. Clin. Investig. 2018, 48 (Suppl. S2), e12943. [Google Scholar] [CrossRef] [PubMed]
- Gajendran, C.; Fukui, S.; Sadhu, N.M.; Zainuddin, M.; Rajagopal, S.; Gosu, R.; Gutch, S.; Fukui, S.; Sheehy, C.E.; Chu, L.; et al. Alleviation of arthritis through prevention of neutrophil extracellular traps by an orally available inhibitor of protein arginine deiminase 4. Sci. Rep. 2023, 13, 3189. [Google Scholar] [CrossRef]
- Pillay, J.; Hietbrink, F.; Koenderman, L.; Leenen, L.P. The systemic inflammatory response induced by trauma is reflected by multiple phenotypes of blood neutrophils. Injury 2007, 38, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Soler-Rodriguez, A.M.; Zhang, H.; Lichenstein, H.S.; Qureshi, N.; Niesel, D.W.; Crowe, S.E.; Peterson, J.W.; Klimpel, G.R. Neutrophil activation by bacterial lipoprotein versus lipopolysaccharide: Differential requirements for serum and CD14. J. Immunol. 2000, 164, 2674–2683. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Hongu, T.; Sakamoto, M.; Funakoshi, Y.; Kanaho, Y. Molecular mechanisms of N-formyl-methionyl-leucyl-phenylalanine-induced superoxide generation and degranulation in mouse neutrophils: Phospholipase D is dispensable. Mol. Cell. Biol. 2013, 33, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, F.; Parisi, M.; Ammendola, R. Distinct signaling cascades elicited by different formyl peptide receptor 2 (FPR2) agonists. Int. J. Mol. Sci. 2013, 14, 7193–7230. [Google Scholar] [CrossRef] [PubMed]
- Metzemaekers, M.; Gouwy, M.; Proost, P. Neutrophil chemoattractant receptors in health and disease: Double-edged swords. Cell. Mol. Immunol. 2020, 17, 433–450. [Google Scholar] [CrossRef]
- Sun, L.; Ye, R.D. Role of G protein-coupled receptors in inflammation. Acta Pharmacol. Sin. 2012, 33, 342–350. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Bauer, A.T.; Kirschfink, M.; Ding, P.; Gebhardt, C.; Borsig, L.; Tüting, T.; Renné, T.; Häffner, K.; et al. Neutrophils activated by membrane attack complexes increase the permeability of melanoma blood vessels. Proc. Natl. Acad. Sci. USA 2022, 119, e2122716119. [Google Scholar] [CrossRef]
- Wang, Y.; Jönsson, F. Expression, Role, and Regulation of Neutrophil Fcγ Receptors. Front. Immunol. 2019, 10, 1958. [Google Scholar] [CrossRef]
- Ben Mkaddem, S.; Benhamou, M.; Monteiro, R.C. Understanding Fc Receptor Involvement in Inflammatory Diseases: From Mechanisms to New Therapeutic Tools. Front. Immunol. 2019, 10, 811. [Google Scholar] [CrossRef] [PubMed]
- Futosi, K.; Fodor, S.; Mócsai, A. Neutrophil cell surface receptors and their intracellular signal transduction pathways. Int. Immunopharmacol. 2013, 17, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Futosi, K.; Mócsai, A. Tyrosine kinase signaling pathways in neutrophils. Immunol. Rev. 2016, 273, 121–139. [Google Scholar] [CrossRef] [PubMed]
- Yago, T.; Shao, B.; Miner, J.J.; Yao, L.; Klopocki, A.G.; Maeda, K.; Coggeshall, K.M.; McEver, R.P. E-selectin engages PSGL-1 and CD44 through a common signaling pathway to induce integrin alphaLbeta2-mediated slow leukocyte rolling. Blood 2010, 116, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Huang, B.; Pan, Y.; Fang, J.; Wang, H.; Ji, Y.; Ling, Y.; Guo, P.; Lin, J.; Li, Q.; et al. Spatiotemporal characteristics of P-selectin-induced β(2) integrin activation of human neutrophils under flow. Front. Immunol. 2022, 13, 1023865. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.J.; Schroder, K. Pattern recognition receptor function in neutrophils. Trends Immunol. 2013, 34, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal Transduct. Target. Ther. 2021, 6, 291. [Google Scholar] [CrossRef]
- De Oliveira, S.; Rosowski, E.E.; Huttenlocher, A. Neutrophil migration in infection and wound repair: Going forward in reverse. Nat. Rev. Immunol. 2016, 16, 378–391. [Google Scholar] [CrossRef]
- Petri, B.; Sanz, M.J. Neutrophil chemotaxis. Cell Tissue Res. 2018, 371, 425–436. [Google Scholar] [CrossRef]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef]
- Segal, A.W. How neutrophils kill microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef] [PubMed]
- Liew, P.X.; Kubes, P. The Neutrophil’s Role During Health and Disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.R.; Robb, C.T.; Perretti, M.; Rossi, A.G. The role of neutrophils in inflammation resolution. Semin. Immunol. 2016, 28, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.F.; Cao, K.; Jiang, J.P.; Guan, W.X.; Du, J.F. Neutrophil dysregulation during sepsis: An overview and update. J. Cell. Mol. Med. 2017, 21, 1687–1697. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kaplan, M.J. Neutrophil Dysregulation in the Pathogenesis of Systemic Lupus Erythematosus. Rheum. Dis. Clin. North Am. 2021, 47, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, A.; Herrmann, M.; Muñoz, L.E. Clearance Deficiency and Cell Death Pathways: A Model for the Pathogenesis of SLE. Front. Immunol. 2016, 7, 35. [Google Scholar] [CrossRef] [PubMed]
- Schuster, S.; Hurrell, B.; Tacchini-Cottier, F. Crosstalk between neutrophils and dendritic cells: A context-dependent process. J. Leukoc. Biol. 2013, 94, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Prame Kumar, K.; Nicholls, A.J.; Wong, C.H.Y. Partners in crime: Neutrophils and monocytes/macrophages in inflammation and disease. Cell Tissue Res. 2018, 371, 551–565. [Google Scholar] [CrossRef]
- Soehnlein, O.; Zernecke, A.; Eriksson, E.E.; Rothfuchs, A.G.; Pham, C.T.; Herwald, H.; Bidzhekov, K.; Rottenberg, M.E.; Weber, C.; Lindbom, L. Neutrophil secretion products pave the way for inflammatory monocytes. Blood 2008, 112, 1461–1471. [Google Scholar] [CrossRef]
- Soehnlein, O.; Lindbom, L.; Weber, C. Mechanisms underlying neutrophil-mediated monocyte recruitment. Blood 2009, 114, 4613–4623. [Google Scholar] [CrossRef]
- Soehnlein, O.; Weber, C.; Lindbom, L. Neutrophil granule proteins tune monocytic cell function. Trends Immunol. 2009, 30, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Nahrendorf, M.; Swirski, F.K. Neutrophil-macrophage communication in inflammation and atherosclerosis. Science 2015, 349, 237–238. [Google Scholar] [CrossRef] [PubMed]
- Scapini, P.; Bazzoni, F.; Cassatella, M.A. Regulation of B-cell-activating factor (BAFF)/B lymphocyte stimulator (BLyS) expression in human neutrophils. Immunol. Lett. 2008, 116, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Bevilacqua, D.; Cassatella, M.A.; Scapini, P. Recent advances on the crosstalk between neutrophils and B or T lymphocytes. Immunology 2019, 156, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Tate, M.D.; Brooks, A.G.; Reading, P.C.; Mintern, J.D. Neutrophils sustain effective CD8(+) T-cell responses in the respiratory tract following influenza infection. Immunol. Cell Biol. 2012, 90, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Minns, D.; Smith, K.J.; Hardisty, G.; Rossi, A.G.; Gwyer Findlay, E. The Outcome of Neutrophil-T Cell Contact Differs Depending on Activation Status of Both Cell Types. Front. Immunol. 2021, 12, 633486. [Google Scholar] [CrossRef] [PubMed]
- Abi Abdallah, D.S.; Egan, C.E.; Butcher, B.A.; Denkers, E.Y. Mouse neutrophils are professional antigen-presenting cells programmed to instruct Th1 and Th17 T-cell differentiation. Int. Immunol. 2011, 23, 317–326. [Google Scholar] [CrossRef]
- Minns, D.; Smith, K.J.; Alessandrini, V.; Hardisty, G.; Melrose, L.; Jackson-Jones, L.; MacDonald, A.S.; Davidson, D.J.; Gwyer Findlay, E. The neutrophil antimicrobial peptide cathelicidin promotes Th17 differentiation. Nat. Commun. 2021, 12, 1285. [Google Scholar] [CrossRef]
- Costantini, C.; Cassatella, M.A. The defensive alliance between neutrophils and NK cells as a novel arm of innate immunity. J. Leukoc. Biol. 2011, 89, 221–233. [Google Scholar] [CrossRef]
- Witter, A.R.; Okunnu, B.M.; Berg, R.E. The Essential Role of Neutrophils during Infection with the Intracellular Bacterial Pathogen Listeria monocytogenes. J. Immunol. 2016, 197, 1557–1565. [Google Scholar] [CrossRef]
- Liu, M.; Chen, K.; Yoshimura, T.; Liu, Y.; Gong, W.; Wang, A.; Gao, J.L.; Murphy, P.M.; Wang, J.M. Formylpeptide receptors are critical for rapid neutrophil mobilization in host defense against Listeria monocytogenes. Sci. Rep. 2012, 2, 786. [Google Scholar] [CrossRef]
- Golenkina, E.A.; Galkina, S.I.; Pletjushkina, O.; Chernyak, B.; Gaponova, T.V.; Romanova, Y.M.; Sud’ina, G.F. Gram-Negative Bacteria Salmonella typhimurium Boost Leukotriene Synthesis Induced by Chemoattractant fMLP to Stimulate Neutrophil Swarming. Front. Pharmacol. 2021, 12, 814113. [Google Scholar] [CrossRef]
- Lin, A.; Loughman, J.A.; Zinselmeyer, B.H.; Miller, M.J.; Caparon, M.G. Streptolysin S inhibits neutrophil recruitment during the early stages of Streptococcus pyogenes infection. Infect. Immun. 2009, 77, 5190–5201. [Google Scholar] [CrossRef] [PubMed]
- Surewaard, B.G.; Trzciński, K.; Jacobino, S.R.; Hansen, I.S.; Vughs, M.M.; Sanders, E.A.; van der Ende, A.; van Strijp, J.A.; de Haas, C.J. Pneumococcal immune evasion: ZmpC inhibits neutrophil influx. Cell. Microbiol. 2013, 15, 1753–1765. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, M.; Heinzinger, L.R.; Morenikeji, O.B.; Marzullo, B.; Thomas, B.N.; Bou Ghanem, E.N. Transcriptome Profiling Reveals CD73 and Age-Driven Changes in Neutrophil Responses against Streptococcus pneumoniae. Infect. Immun. 2021, 89, e0025821. [Google Scholar] [CrossRef] [PubMed]
- Spaan, A.N.; Surewaard, B.G.; Nijland, R.; van Strijp, J.A. Neutrophils versus Staphylococcus aureus: A biological tug of war. Annu. Rev. Microbiol. 2013, 67, 629–650. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.D.; Malachowa, N.; DeLeo, F.R. Neutrophils and Bacterial Immune Evasion. J. Innate Immun. 2018, 10, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.; Teghanemt, A.; Apicella, M.A.; Nauseef, W.M. Modulation of phagocytosis-induced cell death of human neutrophils by Neisseria gonorrhoeae. J. Leukoc. Biol. 2020, 108, 1543–1553. [Google Scholar] [CrossRef] [PubMed]
- Hyams, C.; Camberlein, E.; Cohen, J.M.; Bax, K.; Brown, J.S. The Streptococcus pneumoniae capsule inhibits complement activity and neutrophil phagocytosis by multiple mechanisms. Infect. Immun. 2010, 78, 704–715. [Google Scholar] [CrossRef]
- Ninkovic, J.; Anand, V.; Dutta, R.; Zhang, L.; Saluja, A.; Meng, J.; Koodie, L.; Banerjee, S.; Roy, S. Differential effects of gram-positive and gram-negative bacterial products on morphine induced inhibition of phagocytosis. Sci. Rep. 2016, 6, 21094. [Google Scholar] [CrossRef]
- Parker, H.A.; Forrester, L.; Kaldor, C.D.; Dickerhof, N.; Hampton, M.B. Antimicrobial Activity of Neutrophils Against Mycobacteria. Front. Immunol. 2021, 12, 782495. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.D.; Porter, A.R.; Dorward, D.W.; Brinkworth, A.J.; Chen, L.; Kreiswirth, B.N.; DeLeo, F.R. Phagocytosis and Killing of Carbapenem-Resistant ST258 Klebsiella pneumoniae by Human Neutrophils. J. Infect. Dis. 2016, 213, 1615–1622. [Google Scholar] [CrossRef] [PubMed]
- Managò, A.; Becker, K.A.; Carpinteiro, A.; Wilker, B.; Soddemann, M.; Seitz, A.P.; Edwards, M.J.; Grassmé, H.; Szabò, I.; Gulbins, E. Pseudomonas aeruginosa pyocyanin induces neutrophil death via mitochondrial reactive oxygen species and mitochondrial acid sphingomyelinase. Antioxid. Redox Signal. 2015, 22, 1097–1110. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas aeruginosa: Pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Figueroa, E.; Álvarez-Carrasco, P.; Ortega, E.; Maldonado-Bernal, C. Neutrophils: Many Ways to Die. Front. Immunol. 2021, 12, 631821. [Google Scholar] [CrossRef] [PubMed]
- Demeyere, K.; Remijsen, Q.; Demon, D.; Breyne, K.; Notebaert, S.; Boyen, F.; Guérin, C.J.; Vandenabeele, P.; Meyer, E. Escherichia coli induces bovine neutrophil cell death independent from caspase-3/-7/-1, but with phosphatidylserine exposure prior to membrane rupture. Vet. Immunol. Immunopathol. 2013, 153, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Jiang, C.; Yang, M.; Xiao, H.; Huang, X.; Wu, L.; Yao, M. Salmonella enterica serovar Typhimurium inhibits the innate immune response and promotes apoptosis in a ribosomal/TRP53-dependent manner in swine neutrophils. Vet. Res. 2020, 51, 105. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Delgado-Rizo, V.; Martínez-Guzmán, M.A.; Iñiguez-Gutierrez, L.; García-Orozco, A.; Alvarado-Navarro, A.; Fafutis-Morris, M. Neutrophil Extracellular Traps and Its Implications in Inflammation: An Overview. Front. Immunol. 2017, 8, 81. [Google Scholar] [CrossRef]
- Hung, S.L.; Chiang, H.H.; Wu, C.Y.; Hsu, M.J.; Chen, Y.T. Effects of herpes simplex virus type 1 infection on immune functions of human neutrophils. J. Periodontal Res. 2012, 47, 635–644. [Google Scholar] [CrossRef]
- Stokes, K.L.; Currier, M.G.; Sakamoto, K.; Lee, S.; Collins, P.L.; Plemper, R.K.; Moore, M.L. The respiratory syncytial virus fusion protein and neutrophils mediate the airway mucin response to pathogenic respiratory syncytial virus infection. J. Virol. 2013, 87, 10070–10082. [Google Scholar] [CrossRef] [PubMed]
- Hartshorn, K.L. Innate Immunity and Influenza A Virus Pathogenesis: Lessons for COVID-19. Front. Cell. Infect. Microbiol. 2020, 10, 563850. [Google Scholar] [CrossRef] [PubMed]
- Hensley-McBain, T.; Klatt, N.R. The Dual Role of Neutrophils in HIV Infection. Curr. HIV/AIDS Rep. 2018, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cloke, T.; Munder, M.; Taylor, G.; Müller, I.; Kropf, P. Characterization of a novel population of low-density granulocytes associated with disease severity in HIV-1 infection. PLoS ONE 2012, 7, e48939. [Google Scholar] [CrossRef] [PubMed]
- Leber, B.; Balázs, I.; Horvath, A.; Posch, A.; Streit, A.; Spindelböck, W.; Feldbacher, N.; Stiegler, P.; Stauber, R.E.; Rechberger, G.N.; et al. Direct acting antiviral therapy rescues neutrophil dysfunction and reduces hemolysis in hepatitis C infection. Transl. Res. 2021, 232, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.C.; Weston-Davies, W.; Russo, R.C.; Tavares, L.P.; Rachid, M.A.; Alves-Filho, J.C.; Machado, A.V.; Ryffel, B.; Nunn, M.A.; Teixeira, M.M. Complement C5 activation during influenza A infection in mice contributes to neutrophil recruitment and lung injury. PLoS ONE 2013, 8, e64443. [Google Scholar] [CrossRef] [PubMed]
- Othumpangat, S.; Beezhold, D.H.; Noti, J.D. Influenza virus infection modulates the death receptor pathway during early stages of infection in human bronchial epithelial cells. Physiol. Genom. 2018, 50, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Salmen, S.; Montes, H.; Soyano, A.; Hernández, D.; Berrueta, L. Mechanisms of neutrophil death in human immunodeficiency virus-infected patients: Role of reactive oxygen species, caspases and map kinase pathways. Clin. Exp. Immunol. 2007, 150, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Elbim, C.; Monceaux, V.; François, S.; Hurtrel, B.; Gougerot-Pocidalo, M.A.; Estaquier, J. Increased neutrophil apoptosis in chronically SIV-infected macaques. Retrovirology 2009, 6, 29. [Google Scholar] [CrossRef]
- Rawat, S.; Vrati, S.; Banerjee, A. Neutrophils at the crossroads of acute viral infections and severity. Mol. Asp. Med. 2021, 81, 100996. [Google Scholar] [CrossRef]
- Callejas Caballero, I.; Illán Ramos, M.; Berzosa Sánchez, A.; Anguita, E.; Ramos Amador, J.T. Autoimmune neutropenia associated with influenza virus infection in childhood: A case report. BMC Infect. Dis. 2021, 21, 830. [Google Scholar] [CrossRef] [PubMed]
- Leroi, C.; Balestre, E.; Messou, E.; Minga, A.; Sawadogo, A.; Drabo, J.; Maiga, M.; Zannou, M.; Seydi, M.; Dabis, F.; et al. Incidence of Severe Neutropenia in HIV-Infected People Starting Antiretroviral Therapy in West Africa. PLoS ONE 2017, 12, e0170753. [Google Scholar] [CrossRef] [PubMed]
- Pocock, J.M.; Storisteanu, D.M.L.; Reeves, M.B.; Juss, J.K.; Wills, M.R.; Cowburn, A.S.; Chilvers, E.R. Human Cytomegalovirus Delays Neutrophil Apoptosis and Stimulates the Release of a Prosurvival Secretome. Front. Immunol. 2017, 8, 1185. [Google Scholar] [CrossRef] [PubMed]
- Sebina, I.; Phipps, S. The Contribution of Neutrophils to the Pathogenesis of RSV Bronchiolitis. Viruses 2020, 12, 808. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Komano, J.; Saitoh, Y.; Misawa, T.; Takahama, M.; Kozaki, T.; Uehata, T.; Iwasaki, H.; Omori, H.; Yamaoka, S.; et al. Neutrophil extracellular traps mediate a host defense response to human immunodeficiency virus-1. Cell Host Microbe 2012, 12, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Narasaraju, T.; Yang, E.; Samy, R.P.; Ng, H.H.; Poh, W.P.; Liew, A.A.; Phoon, M.C.; van Rooijen, N.; Chow, V.T. Excessive neutrophils and neutrophil extracellular traps contribute to acute lung injury of influenza pneumonitis. Am. J. Pathol. 2011, 179, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Muraro, S.P.; De Souza, G.F.; Gallo, S.W.; Da Silva, B.K.; De Oliveira, S.D.; Vinolo, M.A.R.; Saraiva, E.M.; Porto, B.N. Respiratory Syncytial Virus induces the classical ROS-dependent NETosis through PAD-4 and necroptosis pathways activation. Sci. Rep. 2018, 8, 14166. [Google Scholar] [CrossRef]
- Diniz, L.F.A.; Matsuba, B.K.; Souza, P.S.S.; Lopes, B.R.P.; Kubo, L.H.; Oliveira, J.; Toledo, K.A. Effects of neutrophil extracellular traps during human respiratory syncytial virus infection in vitro. Braz. J. Biol. 2021, 83, e248717. [Google Scholar] [CrossRef]
- Mutua, V.; Gershwin, L.J. A Review of Neutrophil Extracellular Traps (NETs) in Disease: Potential Anti-NETs Therapeutics. Clin. Rev. Allergy Immunol. 2021, 61, 194–211. [Google Scholar] [CrossRef]
- Othman, A.; Sekheri, M.; Filep, J.G. Roles of neutrophil granule proteins in orchestrating inflammation and immunity. FEBS J. 2022, 289, 3932–3953. [Google Scholar] [CrossRef]
- Filippi, M.; Bar-Or, A.; Piehl, F.; Preziosa, P.; Solari, A.; Vukusic, S.; Rocca, M.A. Multiple sclerosis. Nat. Rev. Dis. Primers 2018, 4, 43. [Google Scholar] [CrossRef]
- De Bondt, M.; Hellings, N.; Opdenakker, G.; Struyf, S. Neutrophils: Underestimated Players in the Pathogenesis of Multiple Sclerosis (MS). Int. J. Mol. Sci. 2020, 21, 4558. [Google Scholar] [CrossRef] [PubMed]
- Akaishi, T.; Misu, T.; Fujihara, K.; Nakaya, N.; Nakamura, T.; Kogure, M.; Hatanaka, R.; Itabashi, F.; Kanno, I.; Takahashi, T.; et al. White blood cell count profiles in multiple sclerosis during attacks before the initiation of acute and chronic treatments. Sci. Rep. 2021, 11, 22357. [Google Scholar] [CrossRef] [PubMed]
- Rossi, B.; Santos-Lima, B.; Terrabuio, E.; Zenaro, E.; Constantin, G. Common Peripheral Immunity Mechanisms in Multiple Sclerosis and Alzheimer’s Disease. Front. Immunol. 2021, 12, 639369. [Google Scholar] [CrossRef] [PubMed]
- International Multiple Sclerosis Genetics Consortium. Multiple sclerosis genomic map implicates peripheral immune cells and microglia in susceptibility. Science 2019, 365, eaav7188. [Google Scholar] [CrossRef] [PubMed]
- Rumble, J.M.; Huber, A.K.; Krishnamoorthy, G.; Srinivasan, A.; Giles, D.A.; Zhang, X.; Wang, L.; Segal, B.M. Neutrophil-related factors as biomarkers in EAE and MS. J. Exp. Med. 2015, 212, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.; Thomas, T.L.; Betmouni, S.; Scolding, N.; Love, S. Elevated myeloperoxidase activity in white matter in multiple sclerosis. Neurosci. Lett. 2008, 444, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.; Thomas, T.L.; Betmouni, S.; Scolding, N.; Love, S. Elevated activity and microglial expression of myeloperoxidase in demyelinated cerebral cortex in multiple sclerosis. Brain Pathol. 2008, 18, 86–95. [Google Scholar] [CrossRef]
- George, J.C.; Mohan, P.; Ho, K. Antineutrophil Cytoplasmic Antibody and Multiple Sclerosis. Kidney Int. Rep. 2018, 3, 732–737. [Google Scholar] [CrossRef]
- Burster, T.; Beck, A.; Tolosa, E.; Marin-Esteban, V.; Rötzschke, O.; Falk, K.; Lautwein, A.; Reich, M.; Brandenburg, J.; Schwarz, G.; et al. Cathepsin G, and not the asparagine-specific endoprotease, controls the processing of myelin basic protein in lysosomes from human B lymphocytes. J. Immunol. 2004, 172, 5495–5503. [Google Scholar] [CrossRef]
- Gao, S.; Zhu, H.; Zuo, X.; Luo, H. Cathepsin G and Its Role in Inflammation and Autoimmune Diseases. Arch. Rheumatol. 2018, 33, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, A.; Nam, Y.; Seo, D.; Rahman, M.H.; Hwang, E.M.; Kim, S.C.; Lee, W.H.; Suk, K. Cathelicidin-related antimicrobial peptide promotes neuroinflammation through astrocyte-microglia communication in experimental autoimmune encephalomyelitis. Glia 2022, 70, 1902–1926. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.J.; Minns, D.; McHugh, B.J.; Holloway, R.K.; O’Connor, R.; Williams, A.; Melrose, L.; McPherson, R.; Miron, V.E.; Davidson, D.J.; et al. The antimicrobial peptide cathelicidin drives development of experimental autoimmune encephalomyelitis in mice by affecting Th17 differentiation. PLoS Biol. 2022, 20, e3001554. [Google Scholar] [CrossRef] [PubMed]
- Al Nimer, F.; Elliott, C.; Bergman, J.; Khademi, M.; Dring, A.M.; Aeinehband, S.; Bergenheim, T.; Romme Christensen, J.; Sellebjerg, F.; Svenningsson, A.; et al. Lipocalin-2 is increased in progressive multiple sclerosis and inhibits remyelination. Neurol. Neuroimmunol. Neuroinflamm. 2016, 3, e191. [Google Scholar] [CrossRef] [PubMed]
- Gasterich, N.; Bohn, A.; Sesterhenn, A.; Nebelo, F.; Fein, L.; Kaddatz, H.; Nyamoya, S.; Kant, S.; Kipp, M.; Weiskirchen, R.; et al. Lipocalin 2 attenuates oligodendrocyte loss and immune cell infiltration in mouse models for multiple sclerosis. Glia 2022, 70, 2188–2206. [Google Scholar] [CrossRef] [PubMed]
- Aubé, B.; Lévesque, S.A.; Paré, A.; Chamma, É.; Kébir, H.; Gorina, R.; Lécuyer, M.A.; Alvarez, J.I.; De Koninck, Y.; Engelhardt, B.; et al. Neutrophils mediate blood-spinal cord barrier disruption in demyelinating neuroinflammatory diseases. J. Immunol. 2014, 193, 2438–2454. [Google Scholar] [CrossRef] [PubMed]
- Brkic, M.; Balusu, S.; Libert, C.; Vandenbroucke, R.E. Friends or Foes: Matrix Metalloproteinases and Their Multifaceted Roles in Neurodegenerative Diseases. Mediat. Inflamm. 2015, 2015, 620581. [Google Scholar] [CrossRef]
- Ohl, K.; Tenbrock, K.; Kipp, M. Oxidative stress in multiple sclerosis: Central and peripheral mode of action. Exp. Neurol. 2016, 277, 58–67. [Google Scholar] [CrossRef]
- Neu, I.S.; Metzger, G.; Zschocke, J.; Zelezny, R.; Mayatepek, E. Leukotrienes in patients with clinically active multiple sclerosis. Acta Neurol. Scand. 2002, 105, 63–66. [Google Scholar] [CrossRef]
- Kihara, Y.; Yokomizo, T.; Kunita, A.; Morishita, Y.; Fukayama, M.; Ishii, S.; Shimizu, T. The leukotriene B4 receptor, BLT1, is required for the induction of experimental autoimmune encephalomyelitis. Biochem. Biophys. Res. Commun. 2010, 394, 673–678. [Google Scholar] [CrossRef]
- He, R.; Chen, Y.; Cai, Q. The role of the LTB4-BLT1 axis in health and disease. Pharmacol. Res. 2020, 158, 104857. [Google Scholar] [CrossRef] [PubMed]
- Olsson, A.; Gustavsen, S.; Hasselbalch, I.C.; Langkilde, A.R.; Sellebjerg, F.; Oturai, A.B.; Søndergaard, H.B. Biomarkers of inflammation and epithelial barrier function in multiple sclerosis. Mult. Scler. Relat. Disord. 2020, 46, 102520. [Google Scholar] [CrossRef] [PubMed]
- Fresegna, D.; Bullitta, S.; Musella, A.; Rizzo, F.R.; De Vito, F.; Guadalupi, L.; Caioli, S.; Balletta, S.; Sanna, K.; Dolcetti, E.; et al. Re-Examining the Role of TNF in MS Pathogenesis and Therapy. Cells 2020, 9, 2290. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Edelson, B.T. New Insights into the Role of IL-1β in Experimental Autoimmune Encephalomyelitis and Multiple Sclerosis. J. Immunol. 2017, 198, 4553–4560. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, C.; Micheletti, A.; Cassatella, M.A. Neutrophil-derived cytokines: Facts beyond expression. Front. Immunol. 2014, 5, 508. [Google Scholar] [CrossRef] [PubMed]
- Grist, J.J.; Marro, B.S.; Skinner, D.D.; Syage, A.R.; Worne, C.; Doty, D.J.; Fujinami, R.S.; Lane, T.E. Induced CNS expression of CXCL1 augments neurologic disease in a murine model of multiple sclerosis via enhanced neutrophil recruitment. Eur. J. Immunol. 2018, 48, 1199–1210. [Google Scholar] [CrossRef]
- Wood, D.D.; Ackerley, C.A.; Brand, B.; Zhang, L.; Raijmakers, R.; Mastronardi, F.G.; Moscarello, M.A. Myelin localization of peptidylarginine deiminases 2 and 4: Comparison of PAD2 and PAD4 activities. Lab. Investig. 2008, 88, 354–364. [Google Scholar] [CrossRef]
- Rohrbach, A.S.; Slade, D.J.; Thompson, P.R.; Mowen, K.A. Activation of PAD4 in NET formation. Front. Immunol. 2012, 3, 360. [Google Scholar] [CrossRef]
- Tillack, K.; Naegele, M.; Haueis, C.; Schippling, S.; Wandinger, K.P.; Martin, R.; Sospedra, M. Gender differences in circulating levels of neutrophil extracellular traps in serum of multiple sclerosis patients. J. Neuroimmunol. 2013, 261, 108–119. [Google Scholar] [CrossRef]
- Brinkman, C.J.; Nillesen, W.M.; Hommes, O.R.; Lamers, K.J.; de Pauw, B.E. Spontaneous and mitogen-induced activity of lymphocytes of different density in multiple sclerosis. Eur. Neurol. 1982, 21, 366–374. [Google Scholar] [CrossRef]
- Ostendorf, L.; Mothes, R.; van Koppen, S.; Lindquist, R.L.; Bellmann-Strobl, J.; Asseyer, S.; Ruprecht, K.; Alexander, T.; Niesner, R.A.; Hauser, A.E.; et al. Low-Density Granulocytes Are a Novel Immunopathological Feature in Both Multiple Sclerosis and Neuromyelitis Optica Spectrum Disorder. Front. Immunol. 2019, 10, 2725. [Google Scholar] [CrossRef] [PubMed]
- Gelibter, S.; Pisa, M.; Croese, T.; Dalla Costa, G.; Orrico, M.; Preziosa, P.; Sangalli, F.; Martinelli, V.; Furlan, R.; Filippi, M. Neutrophil-to-lymphocyte ratio: A marker of neuro-inflammation in multiple sclerosis? J. Neurol. 2021, 268, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Lin, H.C.; Yang, Y.H.; Hsu, C.W.; Chen, N.C.; Tsai, W.C.; Cheng, B.C.; Tsai, N.W. Neutrophil-to-lymphocyte ratio and monocyte-to-lymphocyte ratio are associated with a 2-year relapse in patients with multiple sclerosis. Mult. Scler. Relat. Disord. 2022, 58, 103514. [Google Scholar] [CrossRef]
- Ioannou, M.; Alissafi, T.; Lazaridis, I.; Deraos, G.; Matsoukas, J.; Gravanis, A.; Mastorodemos, V.; Plaitakis, A.; Sharpe, A.; Boumpas, D.; et al. Crucial role of granulocytic myeloid-derived suppressor cells in the regulation of central nervous system autoimmune disease. J. Immunol. 2012, 188, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Geremia, A.; Biancheri, P.; Allan, P.; Corazza, G.R.; Di Sabatino, A. Innate and adaptive immunity in inflammatory bowel disease. Autoimmun. Rev. 2014, 13, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F. Targeting immune cell circuits and trafficking in inflammatory bowel disease. Nat. Immunol. 2019, 20, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.M.; Parkos, C.A. The role of neutrophils during intestinal inflammation. Mucosal Immunol. 2012, 5, 354–366. [Google Scholar] [CrossRef]
- Chami, B.; Martin, N.J.J.; Dennis, J.M.; Witting, P.K. Myeloperoxidase in the inflamed colon: A novel target for treating inflammatory bowel disease. Arch. Biochem. Biophys. 2018, 645, 61–71. [Google Scholar] [CrossRef]
- Hansberry, D.R.; Shah, K.; Agarwal, P.; Agarwal, N. Fecal Myeloperoxidase as a Biomarker for Inflammatory Bowel Disease. Cureus 2017, 9, e1004. [Google Scholar] [CrossRef]
- Curciarello, R.; Sobande, T.; Jones, S.; Giuffrida, P.; Di Sabatino, A.; Docena, G.H.; MacDonald, T.T.; Kok, K. Human Neutrophil Elastase Proteolytic Activity in Ulcerative Colitis Favors the Loss of Function of Therapeutic Monoclonal Antibodies. J. Inflamm. Res 2020, 13, 233–243. [Google Scholar] [CrossRef]
- Kuno, Y.; Ina, K.; Nishiwaki, T.; Tsuzuki, T.; Shimada, M.; Imada, A.; Nishio, Y.; Nobata, K.; Suzuki, T.; Ando, T.; et al. Possible involvement of neutrophil elastase in impaired mucosal repair in patients with ulcerative colitis. J. Gastroenterol. 2002, 37 (Suppl. S14), 22–32. [Google Scholar] [CrossRef]
- Dabek, M.; Ferrier, L.; Roka, R.; Gecse, K.; Annahazi, A.; Moreau, J.; Escourrou, J.; Cartier, C.; Chaumaz, G.; Leveque, M.; et al. Luminal cathepsin g and protease-activated receptor 4: A duet involved in alterations of the colonic epithelial barrier in ulcerative colitis. Am. J. Pathol. 2009, 175, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Zamolodchikova, T.S.; Tolpygo, S.M.; Svirshchevskaya, E.V. Cathepsin G-Not Only Inflammation: The Immune Protease Can Regulate Normal Physiological Processes. Front. Immunol. 2020, 11, 411. [Google Scholar] [CrossRef] [PubMed]
- Owen, C.A.; Campbell, E.J. Angiotensin II generation at the cell surface of activated neutrophils: Novel cathepsin G-mediated catalytic activity that is resistant to inhibition. J. Immunol. 1998, 160, 1436–1443. [Google Scholar] [CrossRef] [PubMed]
- Kanmura, S.; Uto, H.; Numata, M.; Hashimoto, S.; Moriuchi, A.; Fujita, H.; Oketani, M.; Ido, A.; Kodama, M.; Ohi, H.; et al. Human neutrophil peptides 1-3 are useful biomarkers in patients with active ulcerative colitis. Inflamm. Bowel Dis. 2009, 15, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Liu, W.Z.; Zhao, Y.P.; Hu, Y.B.; Ge, Z.Z. Relationship between fecal lactoferrin and inflammatory bowel disease. Scand. J. Gastroenterol. 2007, 42, 1440–1444. [Google Scholar] [CrossRef] [PubMed]
- Krawiec, P.; Pac-Kożuchowska, E. Cathelicidin—A Novel Potential Marker of Pediatric Inflammatory Bowel Disease. J. Inflamm. Res. 2021, 14, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, A.; Holler, E.; Zapater, P.; Sempere, L.; Jover, R.; Pérez-Mateo, M.; Schoelmerich, J.; Such, J.; Wiest, R.; Francés, R. Antimicrobial peptide response to blood translocation of bacterial DNA in Crohn’s disease is affected by NOD2/CARD15 genotype. Inflamm. Bowel Dis. 2011, 17, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, W.; Xiao, W.; Yang, H. The Roles of Cathelicidin LL-37 in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2016, 22, 1986–1991. [Google Scholar] [CrossRef]
- Kusaka, S.; Nishida, A.; Takahashi, K.; Bamba, S.; Yasui, H.; Kawahara, M.; Inatomi, O.; Sugimoto, M.; Andoh, A. Expression of human cathelicidin peptide LL-37 in inflammatory bowel disease. Clin. Exp. Immunol. 2018, 191, 96–106. [Google Scholar] [CrossRef]
- Thorsvik, S.; Damås, J.K.; Granlund, A.V.; Flo, T.H.; Bergh, K.; Østvik, A.E.; Sandvik, A.K. Fecal neutrophil gelatinase-associated lipocalin as a biomarker for inflammatory bowel disease. J. Gastroenterol. Hepatol. 2017, 32, 128–135. [Google Scholar] [CrossRef]
- Vandooren, J.; Van den Steen, P.E.; Opdenakker, G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9): The next decade. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 222–272. [Google Scholar] [CrossRef] [PubMed]
- Mäkitalo, L.; Rintamäki, H.; Tervahartiala, T.; Sorsa, T.; Kolho, K.L. Serum MMPs 7-9 and their inhibitors during glucocorticoid and anti-TNF-α therapy in pediatric inflammatory bowel disease. Scand. J. Gastroenterol. 2012, 47, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, G.; Sipos, F.; Miheller, P.; Hritz, I.; Varga, M.Z.; Juhász, M.; Molnár, B.; Tulassay, Z.; Herszényi, L. The behavior of matrix metalloproteinase-9 in lymphocytic colitis, collagenous colitis and ulcerative colitis. Pathol. Oncol. Res. 2012, 18, 85–91. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.; Gilmer, J.F.; Medina, C. Matrix metalloproteinases in inflammatory bowel disease: An update. Mediat. Inflamm. 2015, 2015, 964131. [Google Scholar] [CrossRef] [PubMed]
- Koelink, P.J.; Overbeek, S.A.; Braber, S.; Morgan, M.E.; Henricks, P.A.; Abdul Roda, M.; Verspaget, H.W.; Wolfkamp, S.C.; te Velde, A.A.; Jones, C.W.; et al. Collagen degradation and neutrophilic infiltration: A vicious circle in inflammatory bowel disease. Gut 2014, 63, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Aviello, G.; Knaus, U.G. ROS in gastrointestinal inflammation: Rescue Or Sabotage? Br. J. Pharmacol 2017, 174, 1704–1718. [Google Scholar] [CrossRef] [PubMed]
- Alzoghaibi, M.A. Concepts of oxidative stress and antioxidant defense in Crohn’s disease. World J. Gastroenterol. 2013, 19, 6540–6547. [Google Scholar] [CrossRef]
- Tian, T.; Wang, Z.; Zhang, J. Pathomechanisms of Oxidative Stress in Inflammatory Bowel Disease and Potential Antioxidant Therapies. Oxidative Med. Cell Longev 2017, 2017, 4535194. [Google Scholar] [CrossRef]
- Sharon, P.; Stenson, W.F. Enhanced synthesis of leukotriene B4 by colonic mucosa in inflammatory bowel disease. Gastroenterology 1984, 86, 453–460. [Google Scholar] [CrossRef]
- Jupp, J.; Hillier, K.; Elliott, D.H.; Fine, D.R.; Bateman, A.C.; Johnson, P.A.; Cazaly, A.M.; Penrose, J.F.; Sampson, A.P. Colonic expression of leukotriene-pathway enzymes in inflammatory bowel diseases. Inflamm. Bowel Dis. 2007, 13, 537–546. [Google Scholar] [CrossRef]
- Lämmermann, T.; Afonso, P.V.; Angermann, B.R.; Wang, J.M.; Kastenmüller, W.; Parent, C.A.; Germain, R.N. Neutrophil swarms require LTB4 and integrins at sites of cell death in vivo. Nature 2013, 498, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, S.; Kuwaki, K.; Mitsuyama, K.; Takedatsu, H.; Yoshioka, S.; Yamasaki, H.; Yamauchi, R.; Mori, A.; Kakuma, T.; Tsuruta, O.; et al. Detection of calprotectin in inflammatory bowel disease: Fecal and serum levels and immunohistochemical localization. Int. J. Mol. Med. 2018, 41, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Shea-Donohue, T.; Thomas, K.; Cody, M.J.; Aiping, Z.; Detolla, L.J.; Kopydlowski, K.M.; Fukata, M.; Lira, S.A.; Vogel, S.N. Mice deficient in the CXCR2 ligand, CXCL1 (KC/GRO-alpha), exhibit increased susceptibility to dextran sodium sulfate (DSS)-induced colitis. Innate Immun. 2008, 14, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, C.; Cassatella, M.A. Neutrophil-derived chemokines on the road to immunity. Semin. Immunol. 2016, 28, 119–128. [Google Scholar] [CrossRef]
- Mouzaoui, S.; Djerdjouri, B.; Makhezer, N.; Kroviarski, Y.; El-Benna, J.; Dang, P.M. Tumor necrosis factor-α-induced colitis increases NADPH oxidase 1 expression, oxidative stress, and neutrophil recruitment in the colon: Preventive effect of apocynin. Mediat. Inflamm. 2014, 2014, 312484. [Google Scholar] [CrossRef]
- Lee, K.M.C.; Achuthan, A.A.; Hamilton, J.A. GM-CSF: A Promising Target in Inflammation and Autoimmunity. Immunotargets Ther. 2020, 9, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wu, X.; Xu, C.; Lin, J.; Liu, Z. Dichotomous roles of neutrophils in modulating pathogenic and repair processes of inflammatory bowel diseases. Precis. Clin. Med. 2021, 4, 246–257. [Google Scholar] [CrossRef]
- Abd El Hafez, A.; Mohamed, A.S.; Shehta, A.; Sheta, H. Neutrophil extracellular traps-associated protein peptidyl arginine deaminase 4 immunohistochemical expression in ulcerative colitis and its association with the prognostic predictors. Pathol. Res. Pract. 2020, 216, 153102. [Google Scholar] [CrossRef]
- Dinallo, V.; Marafini, I.; Di Fusco, D.; Laudisi, F.; Franzè, E.; Di Grazia, A.; Figliuzzi, M.M.; Caprioli, F.; Stolfi, C.; Monteleone, I.; et al. Neutrophil Extracellular Traps Sustain Inflammatory Signals in Ulcerative Colitis. J. Crohns Colitis 2019, 13, 772–784. [Google Scholar] [CrossRef]
- Drury, B.; Hardisty, G.; Gray, R.D.; Ho, G.T. Neutrophil Extracellular Traps in Inflammatory Bowel Disease: Pathogenic Mechanisms and Clinical Translation. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 321–333. [Google Scholar] [CrossRef]
- Acarturk, G.; Acay, A.; Demir, K.; Ulu, M.S.; Ahsen, A.; Yuksel, S. Neutrophil-to-lymphocyte ratio in inflammatory bowel disease—As a new predictor of disease severity. Bratisl. Lek. Listy 2015, 116, 213–217. [Google Scholar] [CrossRef] [PubMed]
- De Cicco, P.; Sanders, T.; Cirino, G.; Maloy, K.J.; Ianaro, A. Hydrogen Sulfide Reduces Myeloid-Derived Suppressor Cell-Mediated Inflammatory Response in a Model of Helicobacter hepaticus-Induced Colitis. Front. Immunol. 2018, 9, 499. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Yu, L.; Fang, L.; Yang, W.; Yu, T.; Miao, Y.; Chen, M.; Wu, K.; Chen, F.; Cong, Y.; et al. CD177(+) neutrophils as functionally activated neutrophils negatively regulate IBD. Gut 2018, 67, 1052–1063. [Google Scholar] [CrossRef]
- Hatemi, G.; Seyahi, E.; Fresko, I.; Talarico, R.; Uçar, D.; Hamuryudan, V. Behçet’s syndrome: One year in review 2022. Clin. Exp. Rheumatol. 2022, 40, 1461–1471. [Google Scholar] [CrossRef] [PubMed]
- Safi, R.; Kallas, R.; Bardawil, T.; Mehanna, C.J.; Abbas, O.; Hamam, R.; Uthman, I.; Kibbi, A.G.; Nassar, D. Neutrophils contribute to vasculitis by increased release of neutrophil extracellular traps in Behçet’s disease. J. Dermatol. Sci. 2018, 92, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Michailidou, D.; Mustelin, T.; Lood, C. Role of Neutrophils in Systemic Vasculitides. Front. Immunol. 2020, 11, 619705. [Google Scholar] [CrossRef] [PubMed]
- Tong, B.; Liu, X.; Xiao, J.; Su, G. Immunopathogenesis of Behcet’s Disease. Front. Immunol. 2019, 10, 665. [Google Scholar] [CrossRef]
- Yu, X.; Li, L.; Zhang, M.; Liu, J.; Chen, H.; Zhang, F.; Zheng, W. Transcriptional analysis of neutrophils from patients with Behçet’s disease reveals activation and chemotaxis of neutrophils. Clin. Immunol. 2022, 245, 109161. [Google Scholar] [CrossRef]
- Ureten, K.; Ertenli, I.; Oztürk, M.A.; Kiraz, S.; Onat, A.M.; Tuncer, M.; Okur, H.; Akdogan, A.; Apras, S.; Calgüneri, M. Neutrophil CD64 expression in Behçet’s disease. J. Rheumatol. 2005, 32, 849–852. [Google Scholar]
- He, K.; Yan, X.; Wu, D. Intestinal Behcet’s Disease: A Review of the Immune Mechanism and Present and Potential Biological Agents. Int. J. Mol. Sci. 2023, 24, 8176. [Google Scholar] [CrossRef]
- Kobayashi, M.; Ito, M.; Nakagawa, A.; Matsushita, M.; Nishikimi, N.; Sakurai, T.; Nimura, Y. Neutrophil and endothelial cell activation in the vasa vasorum in vasculo-Behçet disease. Histopathology 2000, 36, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Pineton de Chambrun, M.; Wechsler, B.; Geri, G.; Cacoub, P.; Saadoun, D. New insights into the pathogenesis of Behçet’s disease. Autoimmun. Rev. 2012, 11, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Le Joncour, A.; Martos, R.; Loyau, S.; Lelay, N.; Dossier, A.; Cazes, A.; Fouret, P.; Domont, F.; Papo, T.; Jandrot-Perrus, M.; et al. Critical role of neutrophil extracellular traps (NETs) in patients with Behcet’s disease. Ann. Rheum. Dis. 2019, 78, 1274–1282. [Google Scholar] [CrossRef]
- Değer, O.; Orem, A.; Akyol, N.; Bahadir, S.; Yildirmis, S. Polymorphonuclear leukocyte elastase levels in patients with Behçet’s disease. Clin. Chim. Acta 1995, 236, 129–134. [Google Scholar] [CrossRef]
- Novak, T.; Fortune, F.; Bergmeier, L.; Khan, I.; Hagi-Pavli, E. Neutrophil elastase and endogenous inhibitors in Behçet’s disease saliva. Clin. Exp. Immunol. 2020, 202, 93–105. [Google Scholar] [CrossRef]
- Kim, M.K.; Kwon, H.C.; Song, J.J.; Park, Y.B.; Lee, S.W. Antineutrophil Cytoplasmic Antibody Positivity Is Associated with Vascular Involvement in Behçet’s Disease. Yonsei Med. J. 2021, 62, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Duzgun, N.; Sahin, M.; Ayaslioglu, E. Anti-Neutrophil Cytoplasmic Antibody in Behçet’s Disease. Int. J. Biomed. Sci. 2006, 2, 49–52. [Google Scholar]
- Brook, M.; Tomlinson, G.H.; Miles, K.; Smith, R.W.; Rossi, A.G.; Hiemstra, P.S.; van ‘t Wout, E.F.; Dean, J.L.; Gray, N.K.; Lu, W.; et al. Neutrophil-derived alpha defensins control inflammation by inhibiting macrophage mRNA translation. Proc. Natl. Acad. Sci. USA 2016, 113, 4350–4355. [Google Scholar] [CrossRef]
- Mumcu, G.; Cimilli, H.; Karacayli, Ü.; Inanc, N.; Türe-Özdemir, F.; Eksioglu-Demiralp, E.; Ergun, T.; Direskeneli, H. Salivary levels of HNP 1-3 are related to oral ulcer activity in Behçet’s disease. Int. J. Dermatol. 2013, 52, 1198–1201. [Google Scholar] [CrossRef]
- Hayashida, M.; Miyoshi, J.; Mitsui, T.; Miura, M.; Saito, D.; Sakuraba, A.; Kawashima, S.; Ikegaya, N.; Fukuoka, K.; Karube, M.; et al. Elevated fecal calprotectin and lactoferrin associated with small intestinal lesions in patients with Behçet disease. J. Gastroenterol. Hepatol. 2020, 35, 1340–1346. [Google Scholar] [CrossRef]
- Jönsson, D.; Nilsson, B.O. The antimicrobial peptide LL-37 is anti-inflammatory and proapoptotic in human periodontal ligament cells. J. Periodontal. Res. 2012, 47, 330–335. [Google Scholar] [CrossRef]
- Si, F.; Lu, Y.; Wen, Y.; Chen, T.; Zhang, Y.; Yang, Y. Cathelicidin (LL-37) causes expression of inflammatory factors in coronary artery endothelial cells of Kawasaki disease by activating TLR4-NF-κB-NLRP3 signaling. Immun. Inflamm. Dis. 2023, 11, e1032. [Google Scholar] [CrossRef]
- Mumcu, G.; Cimilli, H.; Karacayli, U.; Inanc, N.; Ture-Ozdemir, F.; Eksioglu-Demiralp, E.; Ergun, T.; Direskeneli, H. Salivary levels of antimicrobial peptides Hnp 1-3, Ll-37 and S100 in Behcet’s disease. Arch. Oral Biol. 2012, 57, 642–646. [Google Scholar] [CrossRef]
- Kahraman, T.; Gucluler, G.; Simsek, I.; Yagci, F.C.; Yildirim, M.; Ozen, C.; Dinc, A.; Gursel, M.; Ikromzoda, L.; Sutlu, T.; et al. Circulating LL37 targets plasma extracellular vesicles to immune cells and intensifies Behçet’s disease severity. J. Extracell. Vesicles 2017, 6, 1284449. [Google Scholar] [CrossRef]
- Celik, F.; Coteli, E.; Gul, F.C.; Ozsoy, E.; Kobat, S.G.; Karagoz, Z.K.; Aydin, S.; Ugur, K.; Yardim, M.; Sahin, İ.; et al. Interleukin 18, soluble cluster of differentiation 40, platelet factor 4 variant 1, and neutrophil gelatinase-associated lipocalin can be used as biomarkers to aid activity and diagnosis in ocular Behçet’s disease. Int. Ophthalmol. 2022, 42, 3321–3331. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Min, Y.; Park, S.R.; Kim, E.H.; Lee, D.J.; Bang, D.; Lee, E.S. Matrix metalloproteinase-2, -9, -12, and tissue inhibitor of metalloproteinase 2 gene polymorphisms and cutaneous expressions in patients with Behçet’s disease. Tissue Antigens 2012, 79, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Pay, S.; Abbasov, T.; Erdem, H.; Musabak, U.; Simsek, I.; Pekel, A.; Akdogan, A.; Sengul, A.; Dinc, A. Serum MMP-2 and MMP-9 in patients with Behçet’s disease: Do their higher levels correlate to vasculo-Behçet’s disease associated with aneurysm formation? Clin. Exp. Rheumatol. 2007, 25, S70–S75. [Google Scholar]
- Emmi, G.; Becatti, M.; Bettiol, A.; Hatemi, G.; Prisco, D.; Fiorillo, C. Behçet’s Syndrome as a Model of Thrombo-Inflammation: The Role of Neutrophils. Front. Immunol. 2019, 10, 1085. [Google Scholar] [CrossRef]
- Becatti, M.; Emmi, G.; Silvestri, E.; Bruschi, G.; Ciucciarelli, L.; Squatrito, D.; Vaglio, A.; Taddei, N.; Abbate, R.; Emmi, L.; et al. Neutrophil Activation Promotes Fibrinogen Oxidation and Thrombus Formation in Behçet Disease. Circulation 2016, 133, 302–311. [Google Scholar] [CrossRef]
- Li, Z.; Cheng, L.; Zhan, H.; Li, Y. Chemokines and chemokine receptors in Behçet’s disease. Front. Immunol. 2023, 14, 1109147. [Google Scholar] [CrossRef]
- Zhou, Z.Y.; Chen, S.L.; Shen, N.; Lu, Y. Cytokines and Behcet’s disease. Autoimmun. Rev. 2012, 11, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Neves, F.S.; Spiller, F. Possible mechanisms of neutrophil activation in Behçet’s disease. Int. Immunopharmacol. 2013, 17, 1206–1210. [Google Scholar] [CrossRef] [PubMed]
- Verrou, K.M.; Vlachogiannis, N.I.; Ampatziadis-Michailidis, G.; Moulos, P.; Pavlopoulos, G.A.; Hatzis, P.; Kollias, G.; Sfikakis, P.P. Distinct transcriptional profile of blood mononuclear cells in Behçet’s disease: Insights into the central role of neutrophil chemotaxis. Rheumatology 2021, 60, 4910–4919. [Google Scholar] [CrossRef]
- Sayeed, H.M.; Lee, E.S.; Byun, H.O.; Sohn, S. The role of CCR1 and therapeutic effects of anti-CCL3 antibody in herpes simplex virus-induced Behçet’s disease mouse model. Immunology 2019, 158, 206–218. [Google Scholar] [CrossRef]
- Perazzio, S.F.; Soeiro-Pereira, P.V.; Dos Santos, V.C.; de Brito, M.V.; Salu, B.; Oliva, M.L.V.; Stevens, A.M.; de Souza, A.W.S.; Ochs, H.D.; Torgerson, T.R.; et al. Soluble CD40L is associated with increased oxidative burst and neutrophil extracellular trap release in Behçet’s disease. Arthritis Res. Ther. 2017, 19, 235. [Google Scholar] [CrossRef]
- Lee, Y.H.; Song, G.G. Neutrophil-to-lymphocyte ratio, mean platelet volume and platelet-to-lymphocyte ratio in Behçet’s disease and their correlation with disease activity: A meta-analysis. Int. J. Rheum. Dis. 2018, 21, 2180–2187. [Google Scholar] [CrossRef]
- Murad, M.; Low, L.; Davidson, M.; Murray, P.I.; Rauz, S.; Wallace, G.R. Low density neutrophils are increased in patients with Behçet’s disease but do not explain differences in neutrophil function. J. Inflamm. 2022, 19, 5. [Google Scholar] [CrossRef]
- Kim, J.; Kim, B.E.; Leung, D.Y.M. Pathophysiology of atopic dermatitis: Clinical implications. Allergy Asthma Proc. 2019, 40, 84–92. [Google Scholar] [CrossRef]
- Silverberg, J.I. Comorbidities and the impact of atopic dermatitis. Ann. Allergy Asthma Immunol. 2019, 123, 144–151. [Google Scholar] [CrossRef]
- Dhingra, N.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Gittler, J.K.; Shemer, A.; Raz, A.; Fischetti, V.A.; Krueger, J.G.; Guttman-Yassky, E. Attenuated neutrophil axis in atopic dermatitis compared to psoriasis reflects TH17 pathway differences between these diseases. J. Allergy Clin. Immunol. 2013, 132, 498–501.e493. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.M.; Hill, R.Z.; Schwendinger-Schreck, J.; Deguine, J.; Brock, E.C.; Kucirek, N.; Rifi, Z.; Wei, J.; Gronert, K.; Brem, R.B.; et al. Neutrophils promote CXCR3-dependent itch in the development of atopic dermatitis. Elife 2019, 8, e48448. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.L.; Vakharia, P.P.; Silverberg, J.I. The Role and Diagnosis of Allergic Contact Dermatitis in Patients with Atopic Dermatitis. Am. J. Clin. Dermatol. 2018, 19, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Strzepa, A.; Gurski, C.J.; Dittel, L.J.; Szczepanik, M.; Pritchard, K.A., Jr.; Dittel, B.N. Neutrophil-Derived Myeloperoxidase Facilitates Both the Induction and Elicitation Phases of Contact Hypersensitivity. Front. Immunol. 2020, 11, 608871. [Google Scholar] [CrossRef]
- Brunner, P.M.; He, H.; Pavel, A.B.; Czarnowicki, T.; Lefferdink, R.; Erickson, T.; Canter, T.; Puar, N.; Rangel, S.M.; Malik, K.; et al. The blood proteomic signature of early-onset pediatric atopic dermatitis shows systemic inflammation and is distinct from adult long-standing disease. J. Am. Acad. Dermatol. 2019, 81, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Neshkova, E.; Puzhko, S.; Dotsenko, V.; Nenasheva, N.; Yarovaya, G.; Gorjachkina, L. Activity of leukocyte elastase in patients’ plasma is a significant indicator of atopic diseases. Immunopharmacology 1996, 33, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Breuckmann, F.; von Kobyletzki, G.; Avermaete, A.; Kreuter, A.; Altmeyer, P.; Gambichler, T. Modulation of cathepsin G expression in severe atopic dermatitis following medium-dose UVA1 phototherapy. BMC Dermatol. 2002, 2, 12. [Google Scholar] [CrossRef]
- Kim, N.; Bae, K.B.; Kim, M.O.; Yu, D.H.; Kim, H.J.; Yuh, H.S.; Ji, Y.R.; Park, S.J.; Kim, S.; Son, K.H.; et al. Overexpression of cathepsin S induces chronic atopic dermatitis in mice. J. Investig. Dermatol. 2012, 132, 1169–1176. [Google Scholar] [CrossRef]
- Tsybikov, N.N.; Petrisheva, I.V.; Fefelova, E.V.; Kuznik, B.I.; Magen, E. Plasma α-defensins are elevated during exacerbation of atopic dermatitis. Clin. Exp. Dermatol. 2016, 41, 253–259. [Google Scholar] [CrossRef]
- Fujishima, H.; Okada, N.; Matsumoto, K.; Shimizu, E.; Fukuda, S.; Tomita, M. Conjunctival Injection Reduction in Patients with Atopic Keratoconjunctivitis Due to Synergic Effect of Bovine Enteric-Coated Lactoferrin in 0.1% Tacrolimus Ophthalmic Suspension. J. Clin. Med. 2020, 9, 3093. [Google Scholar] [CrossRef]
- Ballardini, N.; Johansson, C.; Lilja, G.; Lindh, M.; Linde, Y.; Scheynius, A.; Agerberth, B. Enhanced expression of the antimicrobial peptide LL-37 in lesional skin of adults with atopic eczema. Br. J. Dermatol. 2009, 161, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Reinholz, M.; Ruzicka, T.; Schauber, J. Cathelicidin LL-37: An antimicrobial peptide with a role in inflammatory skin disease. Ann. Dermatol. 2012, 24, 126–135. [Google Scholar] [CrossRef]
- Nguyen, H.L.T.; Trujillo-Paez, J.V.; Umehara, Y.; Yue, H.; Peng, G.; Kiatsurayanon, C.; Chieosilapatham, P.; Song, P.; Okumura, K.; Ogawa, H.; et al. Role of Antimicrobial Peptides in Skin Barrier Repair in Individuals with Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 7607. [Google Scholar] [CrossRef] [PubMed]
- Suwanchote, S.; Waitayangkoon, P.; Chancheewa, B.; Inthanachai, T.; Niwetbowornchai, N.; Edwards, S.W.; Virakul, S.; Thammahong, A.; Kiatsurayanon, C.; Rerknimitr, P.; et al. Role of antimicrobial peptides in atopic dermatitis. Int. J. Dermatol. 2022, 61, 532–540. [Google Scholar] [CrossRef]
- Choi, J.W.; Lee, M.H.; Fujii, T. Relationship between Neutrophil Gelatinase-Associated Lipocalin, Eosinophil Cationic Protein, Cytokines, and Atopic Sensitization in Patients with Allergic Diseases. Biomed. Res. Int. 2022, 2022, 6564706. [Google Scholar] [CrossRef] [PubMed]
- Bertino, L.; Guarneri, F.; Cannavò, S.P.; Casciaro, M.; Pioggia, G.; Gangemi, S. Oxidative Stress and Atopic Dermatitis. Antioxidants 2020, 9, 196. [Google Scholar] [CrossRef] [PubMed]
- Oyoshi, M.K.; He, R.; Li, Y.; Mondal, S.; Yoon, J.; Afshar, R.; Chen, M.; Lee, D.M.; Luo, H.R.; Luster, A.D.; et al. Leukotriene B4-driven neutrophil recruitment to the skin is essential for allergic skin inflammation. Immunity 2012, 37, 747–758. [Google Scholar] [CrossRef]
- Capucetti, A.; Albano, F.; Bonecchi, R. Multiple Roles for Chemokines in Neutrophil Biology. Front. Immunol. 2020, 11, 1259. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, S.; Farooq, S.M.; Castelvetere, M.P.; Hou, Y.; Gao, J.L.; Navarro, J.V.; Oupicky, D.; Sun, F.; Li, C. A chemokine receptor CXCR2 macromolecular complex regulates neutrophil functions in inflammatory diseases. J. Biol. Chem. 2012, 287, 5744–5755. [Google Scholar] [CrossRef]
- Alam, M.J.; Xie, L.; Ang, C.; Fahimi, F.; Willingham, S.B.; Kueh, A.J.; Herold, M.J.; Mackay, C.R.; Robert, R. Therapeutic blockade of CXCR2 rapidly clears inflammation in arthritis and atopic dermatitis models: Demonstration with surrogate and humanized antibodies. mAbs 2020, 12, 1856460. [Google Scholar] [CrossRef]
- Choy, D.F.; Hsu, D.K.; Seshasayee, D.; Fung, M.A.; Modrusan, Z.; Martin, F.; Liu, F.T.; Arron, J.R. Comparative transcriptomic analyses of atopic dermatitis and psoriasis reveal shared neutrophilic inflammation. J. Allergy Clin. Immunol. 2012, 130, 1335–1343.e1335. [Google Scholar] [CrossRef] [PubMed]
- Geoghegan, J.A.; Irvine, A.D.; Foster, T.J. Staphylococcus aureus and Atopic Dermatitis: A Complex and Evolving Relationship. Trends Microbiol. 2018, 26, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Bitschar, K.; Staudenmaier, L.; Klink, L.; Focken, J.; Sauer, B.; Fehrenbacher, B.; Herster, F.; Bittner, Z.; Bleul, L.; Schaller, M.; et al. Staphylococcus aureus Skin Colonization Is Enhanced by the Interaction of Neutrophil Extracellular Traps with Keratinocytes. J. Investig. Dermatol. 2020, 140, 1054–1065.e1054. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.C.; Yu, H.S.; Yen, F.L.; Lin, C.L.; Chen, G.S.; Lan, C.C. Neutrophil extracellular trap formation is increased in psoriasis and induces human β-defensin-2 production in epidermal keratinocytes. Sci. Rep. 2016, 6, 31119. [Google Scholar] [CrossRef] [PubMed]
- Harder, J.; Dressel, S.; Wittersheim, M.; Cordes, J.; Meyer-Hoffert, U.; Mrowietz, U.; Fölster-Holst, R.; Proksch, E.; Schröder, J.M.; Schwarz, T.; et al. Enhanced expression and secretion of antimicrobial peptides in atopic dermatitis and after superficial skin injury. J. Investig. Dermatol. 2010, 130, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- Batmaz, S.B. Simple Markers for Systemic Inflammation in Pediatric Atopic Dermatitis Patients. Indian J. Dermatol. 2018, 63, 305–310. [Google Scholar] [CrossRef]
- Jiang, Y.; Ma, W. Assessment of Neutrophil-to-Lymphocyte Ratio and Platelet-to-Lymphocyte Ratio in Atopic Dermatitis Patients. Med. Sci. Monit. 2017, 23, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Rafei-Shamsabadi, D.A.; Klose, C.S.N.; Halim, T.Y.F.; Tanriver, Y.; Jakob, T. Context Dependent Role of Type 2 Innate Lymphoid Cells in Allergic Skin Inflammation. Front. Immunol. 2019, 10, 2591. [Google Scholar] [CrossRef]
- Lei, A.; He, Y.; Yang, Q.; Li, X.; Li, R. Role of myeloid cells in the regulation of group 2 innate lymphoid cell-mediated allergic inflammation. Immunology 2020, 161, 18–24. [Google Scholar] [CrossRef]
- Lin, Y.J.; Anzaghe, M.; Schülke, S. Update on the Pathomechanism, Diagnosis, and Treatment Options for Rheumatoid Arthritis. Cells 2020, 9, 880. [Google Scholar] [CrossRef]
- Scherer, H.U.; Häupl, T.; Burmester, G.R. The etiology of rheumatoid arthritis. J. Autoimmun. 2020, 110, 102400. [Google Scholar] [CrossRef]
- Wright, H.L.; Moots, R.J.; Edwards, S.W. The multifactorial role of neutrophils in rheumatoid arthritis. Nat. Rev. Rheumatol. 2014, 10, 593–601. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, L.J.; Kaplan, M.J. Neutrophils in Rheumatoid Arthritis: Breaking Immune Tolerance and Fueling Disease. Trends Mol. Med. 2019, 25, 215–227. [Google Scholar] [CrossRef]
- Wright, H.L.; Lyon, M.; Chapman, E.A.; Moots, R.J.; Edwards, S.W. Rheumatoid Arthritis Synovial Fluid Neutrophils Drive Inflammation Through Production of Chemokines, Reactive Oxygen Species, and Neutrophil Extracellular Traps. Front. Immunol. 2020, 11, 584116. [Google Scholar] [CrossRef]
- Stamp, L.K.; Khalilova, I.; Tarr, J.M.; Senthilmohan, R.; Turner, R.; Haigh, R.C.; Winyard, P.G.; Kettle, A.J. Myeloperoxidase and oxidative stress in rheumatoid arthritis. Rheumatology 2012, 51, 1796–1803. [Google Scholar] [CrossRef]
- Muley, M.M.; Reid, A.R.; Botz, B.; Bölcskei, K.; Helyes, Z.; McDougall, J.J. Neutrophil elastase induces inflammation and pain in mouse knee joints via activation of proteinase-activated receptor-2. Br. J. Pharmacol. 2016, 173, 766–777. [Google Scholar] [CrossRef] [PubMed]
- Nordstrom, D.; Lindy, O.; Konttinen, Y.T.; Lauhio, A.; Sorsa, T.; Friman, C.; Pettersson, T.; Santavirta, S. Cathepsin G and elastase in synovial fluid and peripheral blood in reactive and rheumatoid arthritis. Clin. Rheumatol. 1996, 15, 35–41. [Google Scholar] [CrossRef]
- Behl, T.; Chadha, S.; Sehgal, A.; Singh, S.; Sharma, N.; Kaur, R.; Bhatia, S.; Al-Harrasi, A.; Chigurupati, S.; Alhowail, A.; et al. Exploring the role of cathepsin in rheumatoid arthritis. Saudi J. Biol. Sci. 2022, 29, 402–410. [Google Scholar] [CrossRef]
- Miyata, J.; Tani, K.; Sato, K.; Otsuka, S.; Urata, T.; Lkhagvaa, B.; Furukawa, C.; Sano, N.; Sone, S. Cathepsin G: The significance in rheumatoid arthritis as a monocyte chemoattractant. Rheumatol. Int. 2007, 27, 375–382. [Google Scholar] [CrossRef]
- Okcu, M.; Oktayoglu, P.; Mete, N.; Bozkurt, M.; Caglayan, M.; Dagli, A.Z.; Nas, K. A useful marker in the assessment of remission and activation of disease in patients with rheumatoid arthritis: Serum human neutrophil peptides 1-3. J. Back Musculoskelet. Rehabil. 2018, 31, 1145–1150. [Google Scholar] [CrossRef]
- Umekita, K.; Miyauchi, S.; Nomura, H.; Umeki, K.; Okayama, A. Neutrophil-derived lactoferrin induces the inflammatory responses of rheumatoid arthritis synovial fibroblasts via Toll-like receptor 4. Clin. Exp. Rheumatol. 2019, 37, 834–841. [Google Scholar] [PubMed]
- Cheah, C.W.; Al-Maleki, A.R.; Vadivelu, J.; Danaee, M.; Sockalingam, S.; Baharuddin, N.A.; Vaithilingam, R.D. Salivary and serum cathelicidin LL-37 levels in subjects with rheumatoid arthritis and chronic periodontitis. Int. J. Rheum. Dis. 2020, 23, 1344–1352. [Google Scholar] [CrossRef]
- Hoffmann, M.H.; Bruns, H.; Bäckdahl, L.; Neregård, P.; Niederreiter, B.; Herrmann, M.; Catrina, A.I.; Agerberth, B.; Holmdahl, R. The cathelicidins LL-37 and rCRAMP are associated with pathogenic events of arthritis in humans and rats. Ann. Rheum. Dis. 2013, 72, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Katano, M.; Okamoto, K.; Arito, M.; Kawakami, Y.; Kurokawa, M.S.; Suematsu, N.; Shimada, S.; Nakamura, H.; Xiang, Y.; Masuko, K.; et al. Implication of granulocyte-macrophage colony-stimulating factor induced neutrophil gelatinase-associated lipocalin in pathogenesis of rheumatoid arthritis revealed by proteome analysis. Arthritis Res. Ther. 2009, 11, R3. [Google Scholar] [CrossRef]
- Xue, M.; McKelvey, K.; Shen, K.; Minhas, N.; March, L.; Park, S.Y.; Jackson, C.J. Endogenous MMP-9 and not MMP-2 promotes rheumatoid synovial fibroblast survival, inflammation and cartilage degradation. Rheumatology 2014, 53, 2270–2279. [Google Scholar] [CrossRef]
- Sopata, I.; Wize, J.; Filipowicz-Sosnowska, A.; Stanisławska-Biernat, E.; Brzezińska, B.; Maśliński, S. Neutrophil gelatinase levels in plasma and synovial fluid of patients with rheumatic diseases. Rheumatol. Int. 1995, 15, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Mateen, S.; Moin, S.; Khan, A.Q.; Zafar, A.; Fatima, N. Increased Reactive Oxygen Species Formation and Oxidative Stress in Rheumatoid Arthritis. PLoS ONE 2016, 11, e0152925. [Google Scholar] [CrossRef]
- Sadik, C.D.; Kim, N.D.; Iwakura, Y.; Luster, A.D. Neutrophils orchestrate their own recruitment in murine arthritis through C5aR and FcγR signaling. Proc. Natl. Acad. Sci. USA 2012, 109, E3177–E3185. [Google Scholar] [CrossRef]
- Zheng, L.X.; Li, K.X.; Hong, F.F.; Yang, S.L. Pain and bone damage in rheumatoid arthritis: Role of leukotriene B4. Clin. Exp. Rheumatol. 2019, 37, 872–878. [Google Scholar]
- Miyabe, Y.; Miyabe, C.; Luster, A.D. LTB(4) and BLT1 in inflammatory arthritis. Semin. Immunol. 2017, 33, 52–57. [Google Scholar] [CrossRef]
- Mansour, H.E.; Abdullrhman, M.A.; Mobasher, S.A.; El Mallah, R.; Abaza, N.; Hamed, F.; Khalil, A.A.F. Serum Calprotectin in Rheumatoid Arthritis: A Promising Diagnostic Marker, How Far Is It Related to Activity and Sonographic Findings? J. Med. Ultrasound 2017, 25, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Ometto, F.; Friso, L.; Astorri, D.; Botsios, C.; Raffeiner, B.; Punzi, L.; Doria, A. Calprotectin in rheumatic diseases. Exp. Biol. Med. 2017, 242, 859–873. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chen, W.; Lin, J. The Role of Calprotectin in Rheumatoid Arthritis. J. Transl. Int. Med. 2019, 7, 126–131. [Google Scholar] [CrossRef] [PubMed]
- An, Q.; Yan, W.; Zhao, Y.; Yu, K. Enhanced neutrophil autophagy and increased concentrations of IL-6, IL-8, IL-10 and MCP-1 in rheumatoid arthritis. Int. Immunopharmacol. 2018, 65, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Foulquier, C.; Sebbag, M.; Clavel, C.; Chapuy-Regaud, S.; Al Badine, R.; Méchin, M.C.; Vincent, C.; Nachat, R.; Yamada, M.; Takahara, H.; et al. Peptidyl arginine deiminase type 2 (PAD-2) and PAD-4 but not PAD-1, PAD-3, and PAD-6 are expressed in rheumatoid arthritis synovium in close association with tissue inflammation. Arthritis Rheum. 2007, 56, 3541–3553. [Google Scholar] [CrossRef]
- Spengler, J.; Lugonja, B.; Ytterberg, A.J.; Zubarev, R.A.; Creese, A.J.; Pearson, M.J.; Grant, M.M.; Milward, M.; Lundberg, K.; Buckley, C.D.; et al. Release of Active Peptidyl Arginine Deiminases by Neutrophils Can Explain Production of Extracellular Citrullinated Autoantigens in Rheumatoid Arthritis Synovial Fluid. Arthritis Rheumatol. 2015, 67, 3135–3145. [Google Scholar] [CrossRef] [PubMed]
- Khandpur, R.; Carmona-Rivera, C.; Vivekanandan-Giri, A.; Gizinski, A.; Yalavarthi, S.; Knight, J.S.; Friday, S.; Li, S.; Patel, R.M.; Subramanian, V.; et al. NETs are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci. Transl. Med. 2013, 5, 178ra140. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, C.; Ruiz-Limón, P.; Aguirre, M.A.; Jiménez-Gómez, Y.; Arias-de la Rosa, I.; Ábalos-Aguilera, M.C.; Rodriguez-Ariza, A.; Castro-Villegas, M.C.; Ortega-Castro, R.; Segui, P.; et al. Diagnostic potential of NETosis-derived products for disease activity, atherosclerosis and therapeutic effectiveness in Rheumatoid Arthritis patients. J. Autoimmun. 2017, 82, 31–40. [Google Scholar] [CrossRef]
- Sur Chowdhury, C.; Giaglis, S.; Walker, U.A.; Buser, A.; Hahn, S.; Hasler, P. Enhanced neutrophil extracellular trap generation in rheumatoid arthritis: Analysis of underlying signal transduction pathways and potential diagnostic utility. Arthritis Res. Ther. 2014, 16, R122. [Google Scholar] [CrossRef]
- Liu, X.; Li, J.; Sun, L.; Wang, T.; Liang, W. The association between neutrophil-to-lymphocyte ratio and disease activity in rheumatoid arthritis. Inflammopharmacology 2023, 31, 2237–2244. [Google Scholar] [CrossRef]
- Wright, H.L.; Makki, F.A.; Moots, R.J.; Edwards, S.W. Low-density granulocytes: Functionally distinct, immature neutrophils in rheumatoid arthritis with altered properties and defective TNF signalling. J. Leukoc. Biol. 2017, 101, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Fresneda Alarcon, M.; McLaren, Z.; Wright, H.L. Neutrophils in the Pathogenesis of Rheumatoid Arthritis and Systemic Lupus Erythematosus: Same Foe Different M.O. Front. Immunol. 2021, 12, 649693. [Google Scholar] [CrossRef] [PubMed]
- Pillay, J.; Kamp, V.M.; van Hoffen, E.; Visser, T.; Tak, T.; Lammers, J.W.; Ulfman, L.H.; Leenen, L.P.; Pickkers, P.; Koenderman, L. A subset of neutrophils in human systemic inflammation inhibits T cell responses through Mac-1. J. Clin. Investig. 2012, 122, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Kurkó, J.; Vida, A.; Glant, T.T.; Scanzello, C.R.; Katz, R.S.; Nair, A.; Szekanecz, Z.; Mikecz, K. Identification of myeloid-derived suppressor cells in the synovial fluid of patients with rheumatoid arthritis: A pilot study. BMC Musculoskelet. Disord. 2014, 15, 281. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, S.; Wu, L.; Wang, R.; Zheng, S.; Zhao, D.; Wang, X.; Li, J. The Expansion of Myeloid-Derived Suppressor Cells Is Associated with Joint Inflammation in Rheumatic Patients with Arthritis. Biomed. Res. Int. 2018, 2018, 5474828. [Google Scholar] [CrossRef]
- Wang, W.; Jiao, Z.; Duan, T.; Liu, M.; Zhu, B.; Zhang, Y.; Xu, Q.; Wang, R.; Xiong, Y.; Xu, H.; et al. Functional characterization of myeloid-derived suppressor cell subpopulations during the development of experimental arthritis. Eur. J. Immunol. 2015, 45, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Park, M.J.; Lee, S.H.; Kim, E.K.; Lee, E.J.; Baek, J.A.; Park, S.H.; Kwok, S.K.; Cho, M.L. Interleukin-10 produced by myeloid-derived suppressor cells is critical for the induction of Tregs and attenuation of rheumatoid inflammation in mice. Sci. Rep. 2018, 8, 3753. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhu, D.; Wang, T.; Xia, X.; Tian, J.; Wang, S. Roles of Myeloid-Derived Suppressor Cell Subpopulations in Autoimmune Arthritis. Front. Immunol. 2018, 9, 2849. [Google Scholar] [CrossRef]
- Barber, M.R.W.; Drenkard, C.; Falasinnu, T.; Hoi, A.; Mak, A.; Kow, N.Y.; Svenungsson, E.; Peterson, J.; Clarke, A.E.; Ramsey-Goldman, R. Global epidemiology of systemic lupus erythematosus. Nat. Rev. Rheumatol. 2021, 17, 515–532. [Google Scholar] [CrossRef]
- Rivas-Larrauri, F.; Yamazaki-Nakashimada, M.A. Systemic lupus erythematosus: Is it one disease? Reumatol. Clin. 2016, 12, 274–281. [Google Scholar] [CrossRef]
- Fava, A.; Petri, M. Systemic lupus erythematosus: Diagnosis and clinical management. J. Autoimmun. 2019, 96, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Postal, M.; Vivaldo, J.F.; Fernandez-Ruiz, R.; Paredes, J.L.; Appenzeller, S.; Niewold, T.B. Type I interferon in the pathogenesis of systemic lupus erythematosus. Curr. Opin. Immunol. 2020, 67, 87–94. [Google Scholar] [CrossRef]
- Kaplan, M.J. Neutrophils in the pathogenesis and manifestations of SLE. Nat. Rev. Rheumatol. 2011, 7, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Tsokos, G.C.; Lo, M.S.; Costa Reis, P.; Sullivan, K.E. New insights into the immunopathogenesis of systemic lupus erythematosus. Nat. Rev. Rheumatol. 2016, 12, 716–730. [Google Scholar] [CrossRef]
- Ma, S.; Jiang, W.; Zhang, X.; Liu, W. Insights into the pathogenic role of neutrophils in systemic lupus erythematosus. Curr. Opin. Rheumatol. 2023, 35, 82–88. [Google Scholar] [CrossRef]
- Telles, R.W.; Ferreira, G.A.; da Silva, N.P.; Sato, E.I. Increased plasma myeloperoxidase levels in systemic lupus erythematosus. Rheumatol. Int. 2010, 30, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; Hawkins, C.L. The Role of Myeloperoxidase in Biomolecule Modification, Chronic Inflammation, and Disease. Antioxid. Redox Signal. 2020, 32, 957–981. [Google Scholar] [CrossRef]
- Jeremic, I.; Djuric, O.; Nikolic, M.; Vlajnic, M.; Nikolic, A.; Radojkovic, D.; Bonaci-Nikolic, B. Neutrophil extracellular traps-associated markers are elevated in patients with systemic lupus erythematosus. Rheumatol. Int. 2019, 39, 1849–1857. [Google Scholar] [CrossRef]
- Bruschi, M.; Bonanni, A.; Petretto, A.; Vaglio, A.; Pratesi, F.; Santucci, L.; Migliorini, P.; Bertelli, R.; Galetti, M.; Belletti, S.; et al. Neutrophil Extracellular Traps Profiles in Patients with Incident Systemic Lupus Erythematosus and Lupus Nephritis. J. Rheumatol. 2020, 47, 377–386. [Google Scholar] [CrossRef]
- Tamiya, H.; Tani, K.; Miyata, J.; Sato, K.; Urata, T.; Lkhagvaa, B.; Otsuka, S.; Shigekiyo, S.; Sone, S. Defensins- and cathepsin G-ANCA in systemic lupus erythematosus. Rheumatol. Int. 2006, 27, 147–152. [Google Scholar] [CrossRef]
- Cheng, F.J.; Zhou, X.J.; Zhao, Y.F.; Zhao, M.H.; Zhang, H. Human neutrophil peptide 1-3, a component of the neutrophil extracellular trap, as a potential biomarker of lupus nephritis. Int. J. Rheum. Dis. 2015, 18, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Romo, G.S.; Caielli, S.; Vega, B.; Connolly, J.; Allantaz, F.; Xu, Z.; Punaro, M.; Baisch, J.; Guiducci, C.; Coffman, R.L.; et al. Netting neutrophils are major inducers of type I IFN production in pediatric systemic lupus erythematosus. Sci. Transl. Med. 2011, 3, 73ra20. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Puerta, J.A.; Ortiz-Reyes, B.; Urrego, T.; Vanegas-García, A.L.; Muñoz, C.H.; González, L.A.; Cervera, R.; Vásquez, G. Urinary neutrophil gelatinase-associated lipocalin and monocyte chemoattractant protein 1 as biomarkers for lupus nephritis in Colombian SLE patients. Lupus 2018, 27, 637–646. [Google Scholar] [CrossRef]
- Lindblom, J.; Mohan, C.; Parodis, I. Diagnostic, predictive and prognostic biomarkers in systemic lupus erythematosus: Current insights. Curr. Opin. Rheumatol. 2022, 34, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Faber-Elmann, A.; Sthoeger, Z.; Tcherniack, A.; Dayan, M.; Mozes, E. Activity of matrix metalloproteinase-9 is elevated in sera of patients with systemic lupus erythematosus. Clin. Exp. Immunol. 2002, 127, 393–398. [Google Scholar] [CrossRef]
- Jiang, Z.; Sui, T.; Wang, B. Relationships between MMP-2, MMP-9, TIMP-1 and TIMP-2 levels and their pathogenesis in patients with lupus nephritis. Rheumatol. Int. 2010, 30, 1219–1226. [Google Scholar] [CrossRef]
- Bahrehmand, F.; Vaisi-Raygani, A.; Kiani, A.; Rahimi, Z.; Tavilani, H.; Ardalan, M.; Vaisi-Raygani, H.; Shakiba, E.; Pourmotabbed, T. Matrix metalloproteinase 9 polymorphisms and systemic lupus erythematosus: Correlation with systemic inflammatory markers and oxidative stress. Lupus 2015, 24, 597–605. [Google Scholar] [CrossRef]
- Ehtesham, N.; Mosallaei, M.; Zaboli Mahdiabadi, M.; Kenarangi, T.; Farhadi, A.; Heidari, M.F.; Soroush, M.; Nasrollahzadeh Sabet, M.; Behroozi, J. Significant hypomethylation of MMP9 gene promoter in patients with systemic lupus erythematosus. Lupus 2023, 32, 1056–1065. [Google Scholar] [CrossRef]
- Bengtsson, A.A.; Pettersson, Å.; Wichert, S.; Gullstrand, B.; Hansson, M.; Hellmark, T.; Johansson, Å.C. Low production of reactive oxygen species in granulocytes is associated with organ damage in systemic lupus erythematosus. Arthritis Res. Ther. 2014, 16, R120. [Google Scholar] [CrossRef]
- Wirestam, L.; Arve, S.; Linge, P.; Bengtsson, A.A. Neutrophils-Important Communicators in Systemic Lupus Erythematosus and Antiphospholipid Syndrome. Front. Immunol. 2019, 10, 2734. [Google Scholar] [CrossRef]
- Wu, T.; Xie, C.; Han, J.; Ye, Y.; Weiel, J.; Li, Q.; Blanco, I.; Ahn, C.; Olsen, N.; Putterman, C.; et al. Metabolic disturbances associated with systemic lupus erythematosus. PLoS ONE 2012, 7, e37210. [Google Scholar] [CrossRef] [PubMed]
- Homa-Mlak, I.; Mazurek, M.; Majdan, A.; Mlak, R.; Majdan, M.; Mełecka-Massalska, T. Serum Calprotectin—A NET Product—As a Biomarker of Disease Activity in Patients with Systemic Lupus Erythematosus: A Single-Center Case-Control Study from Poland. Med. Sci. Monit. 2022, 28, e936534. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, M.; Van Hoovels, L.; Benucci, M.; De Luca, R.; Coccia, C.; Bernardini, P.; Russo, E.; Amedei, A.; Guiducci, S.; Grossi, V.; et al. Circulating Calprotectin (cCLP) in autoimmune diseases. Autoimmun. Rev. 2023, 22, 103295. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Pirapakaran, T.; Luo, X.M. Chemokines and Chemokine Receptors in the Development of Lupus Nephritis. Mediat. Inflamm. 2016, 2016, 6012715. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lightfoot, Y.L.; Seto, N.; Carmona-Rivera, C.; Moore, E.; Goel, R.; O’Neil, L.; Mistry, P.; Hoffmann, V.; Mondal, S.; et al. Peptidylarginine deiminases 2 and 4 modulate innate and adaptive immune responses in TLR-7-dependent lupus. JCI Insight 2018, 3, e124729. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Kronbichler, A.; Park, D.D.; Park, Y.; Moon, H.; Kim, H.; Choi, J.H.; Choi, Y.; Shim, S.; Lyu, I.S.; et al. Neutrophil extracellular traps (NETs) in autoimmune diseases: A comprehensive review. Autoimmun. Rev. 2017, 16, 1160–1173. [Google Scholar] [CrossRef] [PubMed]
- Salemme, R.; Peralta, L.N.; Meka, S.H.; Pushpanathan, N.; Alexander, J.J. The Role of NETosis in Systemic Lupus Erythematosus. J. Cell. Immunol. 2019, 1, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Han, B.K.; Wysham, K.D.; Cain, K.C.; Tyden, H.; Bengtsson, A.A.; Lood, C. Neutrophil and lymphocyte counts are associated with different immunopathological mechanisms in systemic lupus erythematosus. Lupus Sci. Med. 2020, 7, e000382. [Google Scholar] [CrossRef]
- Rahman, S.; Sagar, D.; Hanna, R.N.; Lightfoot, Y.L.; Mistry, P.; Smith, C.K.; Manna, Z.; Hasni, S.; Siegel, R.M.; Sanjuan, M.A.; et al. Low-density granulocytes activate T cells and demonstrate a non-suppressive role in systemic lupus erythematosus. Ann. Rheum. Dis. 2019, 78, 957–966. [Google Scholar] [CrossRef]
- Dong, G.; Yang, Y.; Li, X.; Yao, X.; Zhu, Y.; Zhang, H.; Wang, H.; Ma, Q.; Zhang, J.; Shi, H.; et al. Granulocytic myeloid-derived suppressor cells contribute to IFN-I signaling activation of B cells and disease progression through the lncRNA NEAT1-BAFF axis in systemic lupus erythematosus. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165554. [Google Scholar] [CrossRef]
- Abdgawad, M.; Gunnarsson, L.; Bengtsson, A.A.; Geborek, P.; Nilsson, L.; Segelmark, M.; Hellmark, T. Elevated neutrophil membrane expression of proteinase 3 is dependent upon CD177 expression. Clin. Exp. Immunol. 2010, 161, 89–97. [Google Scholar] [CrossRef]
- Bashant, K.R.; Aponte, A.M.; Randazzo, D.; Rezvan Sangsari, P.; Wood, A.J.; Bibby, J.A.; West, E.E.; Vassallo, A.; Manna, Z.G.; Playford, M.P.; et al. Proteomic, biomechanical and functional analyses define neutrophil heterogeneity in systemic lupus erythematosus. Ann. Rheum. Dis. 2021, 80, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Zheng, S.; Zhang, H. Inhibition of myeloperoxidase by N-acetyl lysyltyrosylcysteine amide reduces experimental autoimmune encephalomyelitis-induced injury and promotes oligodendrocyte regeneration and neurogenesis in a murine model of progressive multiple sclerosis. Neuroreport 2018, 29, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Forghani, R.; Wojtkiewicz, G.R.; Zhang, Y.; Seeburg, D.; Bautz, B.R.; Pulli, B.; Milewski, A.R.; Atkinson, W.L.; Iwamoto, Y.; Zhang, E.R.; et al. Demyelinating diseases: Myeloperoxidase as an imaging biomarker and therapeutic target. Radiology 2012, 263, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, G.; Chami, B.; Liu, Y.; Schroder, A.L.; San Gabriel, P.T.; Gao, A.; Fong, G.; Wang, X.; Witting, P.K. The Synthetic Myeloperoxidase Inhibitor AZD3241 Ameliorates Dextran Sodium Sulfate Stimulated Experimental Colitis. Front. Pharmacol. 2020, 11, 556020. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Jiang, H.; Zhu, Q.; Xu, J.; Cai, Y.; Huo, G.; Yuan, K.; Huang, G.; Xu, A. Tetrandrine Ameliorates Rheumatoid Arthritis in Mice by Alleviating Neutrophil Activities. Evid Based Complement. Altern. Med. 2022, 2022, 8589121. [Google Scholar] [CrossRef] [PubMed]
- Norga, K.; Paemen, L.; Masure, S.; Dillen, C.; Heremans, H.; Billiau, A.; Carton, H.; Cuzner, L.; Olsson, T.; Van Damme, J.; et al. Prevention of acute autoimmune encephalomyelitis and abrogation of relapses in murine models of multiple sclerosis by the protease inhibitor D-penicillamine. Inflamm. Res. 1995, 44, 529–534. [Google Scholar] [CrossRef]
- Harper, J.I.; Godwin, H.; Green, A.; Wilkes, L.E.; Holden, N.J.; Moffatt, M.; Cookson, W.O.; Layton, G.; Chandler, S. A study of matrix metalloproteinase expression and activity in atopic dermatitis using a novel skin wash sampling assay for functional biomarker analysis. Br. J. Dermatol. 2010, 162, 397–403. [Google Scholar] [CrossRef]
- Trivedi, P.P.; Jena, G.B. Role of α-lipoic acid in dextran sulfate sodium-induced ulcerative colitis in mice: Studies on inflammation, oxidative stress, DNA damage and fibrosis. Food Chem. Toxicol. 2013, 59, 339–355. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Dunay, I.R.; Fuchs, D.; Trautmann, D.; Fischer, A.; Kühl, A.A.; Loddenkemper, C.; Batra, A.; Siegmund, B.; Krell, H.W.; et al. Selective gelatinase blockage ameliorates acute DSS colitis. Eur. J. Microbiol. Immunol. 2011, 1, 228–236. [Google Scholar] [CrossRef]
- Lesiak, A.; Narbutt, J.; Sysa-Jedrzejowska, A.; Lukamowicz, J.; McCauliffe, D.P.; Wózniacka, A. Effect of chloroquine phosphate treatment on serum MMP-9 and TIMP-1 levels in patients with systemic lupus erythematosus. Lupus 2010, 19, 683–688. [Google Scholar] [CrossRef]
- Jablaoui, A.; Kriaa, A.; Mkaouar, H.; Akermi, N.; Soussou, S.; Wysocka, M.; Wołoszyn, D.; Amouri, A.; Gargouri, A.; Maguin, E.; et al. Fecal Serine Protease Profiling in Inflammatory Bowel Diseases. Front. Cell. Infect. Microbiol. 2020, 10, 21. [Google Scholar] [CrossRef]
- Zhang, T.; Mei, Y.; Dong, W.; Wang, J.; Huang, F.; Wu, J. Evaluation of protein arginine deiminase-4 inhibitor in TNBS- induced colitis in mice. Int. Immunopharmacol. 2020, 84, 106583. [Google Scholar] [CrossRef] [PubMed]
- Knight, J.S.; Zhao, W.; Luo, W.; Subramanian, V.; O’Dell, A.A.; Yalavarthi, S.; Hodgin, J.B.; Eitzman, D.T.; Thompson, P.R.; Kaplan, M.J. Peptidylarginine deiminase inhibition is immunomodulatory and vasculoprotective in murine lupus. J. Clin. Investig. 2013, 123, 2981–2993. [Google Scholar] [CrossRef] [PubMed]
- Chirivi, R.G.S.; van Rosmalen, J.W.G.; van der Linden, M.; Euler, M.; Schmets, G.; Bogatkevich, G.; Kambas, K.; Hahn, J.; Braster, Q.; Soehnlein, O.; et al. Therapeutic ACPA inhibits NET formation: A potential therapy for neutrophil-mediated inflammatory diseases. Cell. Mol. Immunol. 2021, 18, 1528–1544. [Google Scholar] [CrossRef] [PubMed]
- Bettiol, A.; Becatti, M.; Silvestri, E.; Argento, F.R.; Fini, E.; Mannucci, A.; Galora, S.; Mattioli, I.; Urban, M.L.; Malandrino, D.; et al. Neutrophil-mediated mechanisms of damage and in-vitro protective effect of colchicine in non-vascular Behçet’s syndrome. Clin. Exp. Immunol. 2021, 206, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Le Joncour, A.; Régnier, P.; Maciejewski-Duval, A.; Charles, E.; Barete, S.; Fouret, P.; Rosenzwajg, M.; Klatzmann, D.; Cacoub, P.; Saadoun, D. Reduction of Neutrophil Activation by Phosphodiesterase 4 Blockade in Behçet’s Disease. Arthritis Rheumatol. 2023, 75, 1628–1637. [Google Scholar] [CrossRef]
- Yang, F.; Luo, X.; Luo, G.; Zhai, Z.; Zhuang, J.; He, J.; Han, J.; Zhang, Y.; Zhuang, L.; Sun, E.; et al. Inhibition of NET formation by polydatin protects against collagen-induced arthritis. Int. Immunopharmacol. 2019, 77, 105919. [Google Scholar] [CrossRef]
- Ruiz-Limón, P.; Ortega, R.; Arias de la Rosa, I.; Abalos-Aguilera, M.D.C.; Perez-Sanchez, C.; Jimenez-Gomez, Y.; Peralbo-Santaella, E.; Font, P.; Ruiz-Vilches, D.; Ferrin, G.; et al. Tocilizumab improves the proatherothrombotic profile of rheumatoid arthritis patients modulating endothelial dysfunction, NETosis, and inflammation. Transl. Res. 2017, 183, 87–103. [Google Scholar] [CrossRef]
- Huang, G.; Yuan, K.; Zhu, Q.; Zhang, S.; Lu, Q.; Zhu, M.; Sheng, H.; Yu, R.; Luo, G.; Xu, A. Triptolide inhibits the inflammatory activities of neutrophils to ameliorate chronic arthritis. Mol. Immunol. 2018, 101, 210–220. [Google Scholar] [CrossRef]
- Kraaij, T.; Kamerling, S.W.A.; de Rooij, E.N.M.; van Daele, P.L.A.; Bredewold, O.W.; Bakker, J.A.; Bajema, I.M.; Scherer, H.U.; Toes, R.E.M.; Huizinga, T.J.W.; et al. The NET-effect of combining rituximab with belimumab in severe systemic lupus erythematosus. J. Autoimmun. 2018, 91, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Hassanalilou, T.; Khalili, L.; Ghavamzadeh, S.; Shokri, A.; Payahoo, L.; Bishak, Y.K. Role of vitamin D deficiency in systemic lupus erythematosus incidence and aggravation. Autoimmun. Highlights 2017, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.Y.; Kim, J.H.; Kho, A.R.; Kim, I.Y.; Lee, S.H.; Lee, B.E.; Choi, E.; Sohn, M.; Stevenson, M.; Chung, T.N.; et al. Inhibition of NADPH oxidase activation reduces EAE-induced white matter damage in mice. J. Neuroinflamm. 2015, 12, 104. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.H.; Al-Shorbagy, M.Y.; Abdallah, D.M.; Nassar, N.N. Telmisartan attenuates colon inflammation, oxidative perturbations and apoptosis in a rat model of experimental inflammatory bowel disease. PLoS ONE 2014, 9, e97193. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Wan, P.; Zhu, X.D.; Ren, Y.P.; Wang, C.; Yan, R.W.; Guo, Y.; Bai, A.P. Inhibition of NADPH oxidase activities ameliorates DSS-induced colitis. Biochem. Pharmacol. 2018, 158, 126–133. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Z.; Fan, X.; Li, W.; Qu, J.; Dong, C.; Wang, Z.; Ji, Z.; Li, Y. Inhibition of DNM1L and mitochondrial fission attenuates inflammatory response in fibroblast-like synoviocytes of rheumatoid arthritis. J. Cell. Mol. Med. 2020, 24, 1516–1528. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Tian, F.; Zheng, H.; Whitman, S.A.; Lin, Y.; Zhang, Z.; Zhang, N.; Zhang, D.D. Nrf2 suppresses lupus nephritis through inhibition of oxidative injury and the NF-κB-mediated inflammatory response. Kidney Int. 2014, 85, 333–343. [Google Scholar] [CrossRef]
- Herges, K.; de Jong, B.A.; Kolkowitz, I.; Dunn, C.; Mandelbaum, G.; Ko, R.M.; Maini, A.; Han, M.H.; Killestein, J.; Polman, C.; et al. Protective effect of an elastase inhibitor in a neuromyelitis optica-like disease driven by a peptide of myelin oligodendroglial glycoprotein. Mult. Scler. J. 2012, 18, 398–408. [Google Scholar] [CrossRef]
- Shioya, Y.; Katakura, K.; Ohira, H. Neutrophil elastase inhibitor suppresses IL-17 based inflammation of murine experimental colitis. Fukushima J. Med. Sci. 2014, 60, 14–21. [Google Scholar] [CrossRef]
- Di Cesare Mannelli, L.; Micheli, L.; Cinci, L.; Maresca, M.; Vergelli, C.; Pacini, A.; Quinn, M.T.; Paola Giovannoni, M.; Ghelardini, C. Effects of the neutrophil elastase inhibitor EL-17 in rat adjuvant-induced arthritis. Rheumatology 2016, 55, 1285–1294. [Google Scholar] [CrossRef]