


1. Introduction
Toxoplasma gondii infection (TGI) is recognized as a highly prevalent parasitic disease in humans and other warm-blooded animals, especially in tropical and subtropical regions [1,2]. Humans are infected by eating cysts from raw meat (e.g., beef and pork), ingesting oocysts in food contaminated with cat feces, and placental diffusion [3]. TGI may be observed as acute or chronic, with or without signs; however, with the immune system activation, the parasite stays put in tissue cyst forms, gently reproduces host organs, and initiates a latent TGI [4]. Therefore, in immunocompetent persons, TGI is routinely asymptomatic, whereas it is deadly in patients with immune system defects due to reactivation of the infection and subsequent lethal symptoms, e.g., encephalitis [5]. Pyrimethamine (PYM), mainly in combination with sulfadiazine, is still reported as the most effective chemical agent against TGI [6]. This drug is an antagonist of folic acid and can result in the dose-linked suppression of the bone marrow, which is mitigated by parallel administration of folic acid [6]. In addition, there are serious challenges in TGI chemotherapy, such as the inability of common drugs to control and eliminate parasites inside the cyst and eradicate the infection, and the occurrence of adverse side effects, such as osteoporosis, blood poisoning, teratogenicity, and hematuria [7,8,9,10]. Accordingly, it is required to improve the current agents as well as find new agents for the prevention and control of TGI.
Nowadays, interest in using nanotechnology for medical purposes is increasing. Due to having exceptional possessions, such as small size and surface reaction, nanoparticles can be considered as potent candidates in the treatment of numerous diseases, mainly infectious ones [11,12]. Among the metal nanoparticles, copper nanoparticles (CNP) have drawn a lot of interest because of having various biological properties, e.g., antimicrobial, anti-inflammatory, anticancer, and antioxidant [13]. There are different methods for making nanoparticles of various materials with the possibility of controlling their size, composition, and uniformity [14]. The usage of herbal extracts in the phytosynthesis of nanoparticles, which is known as green synthesis, can be used as a nature-friendly method and a suitable alternative to conventional methods such as physical and chemical methods [15]. Lupinus arcticus L. is a plant that belongs to the Fabaceae family and displays various pharmacological effects, such as antidiabetic, anticancer, insecticide, and antifungal effects [16,17]. Despite conducting various studies on the anti-Toxoplasma effects of nanoparticles, different results have been obtained, which is probably due to the synthesis approaches, the parasite strain, and the way they are used [18,19]. The current experimental investigation was planned for the green synthesis, characterization, and efficacy of CNP against T. gondii infection.
2. Results and Discussion
The peak of absorption of CNP acquired from the UV-Vis was observed at 491 nm (Figure 1A). UV-Vis spectroscopy is often used to determine the surface plasmon resonance (SPR) of nanoparticles, which is sensitive to electron oscillations with increased nanoparticle intensity. Due to the increase in surface electrons, the SPR wavelength changes with light absorption in the UV-vis range in metal nanoparticles. Therefore, the appearance of distinct peaks in the known region indicates the formation of nanoparticles. Here, a single peak at 491 nm can be attributed to CNP in the sample solution [20,21].
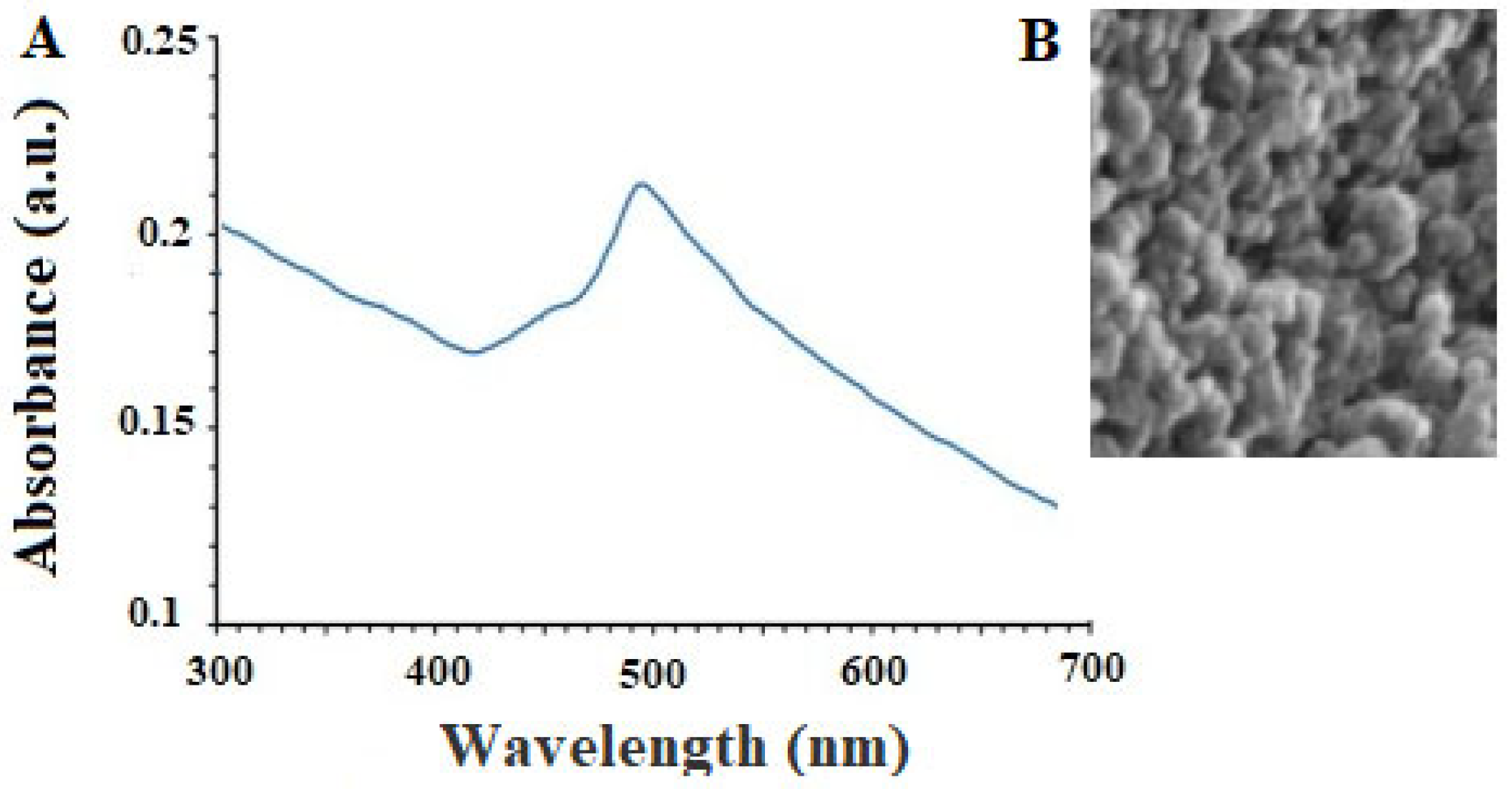
Figure 1. The results of the analysis of UV-Vis (A) and scanning electron microscope (B) of the obtained copper nanoparticles.
SEM analysis also showed that the green synthesized CNP displayed a circular shape with a size ranging from 10 to 85 nm, whereas, the largest spreading of particle size was observed at 30–50 nm (Figure 1B). SEM imaging can be used to determine the nanoparticle morphology and size. Consequently, these two variables are affected by construction conditions, so size and shape can be altered by modifying any environmental factors. Nanoparticles’ stability and biological properties are proportional to their size. Due to their greater surface-to-volume ratio, nanoparticles with a smaller size are more stable and aggregate less frequently. In contrast, coating nanoparticles with molecules can improve their biocompatibility and biological activity. The therapeutic activity of nanoparticles is substantially increased by reducing their size to below 100 nm [22]. This research produced copper nanoparticles smaller than 100 nm, which are suitable for antiparasitic applications [22].
The findings of XRD analysis showed the presence of a diffraction peak at 38.3°, 51.2°, 62.2°, 73.1°, and 83.3° corresponding to (109), (112), (203), (216), and (004), respectively, which represented the monoclinic crystalline phase of CNP (Figure 2A). X-ray diffraction provides data regarding the crystalline phases of the nanoparticles, thereby revealing the nanoparticles’ homogeneity, purity, and crystallite size. In this investigation, the XRD pattern provided specific signals that implied that the purity of the nanoparticles was satisfactory [23].
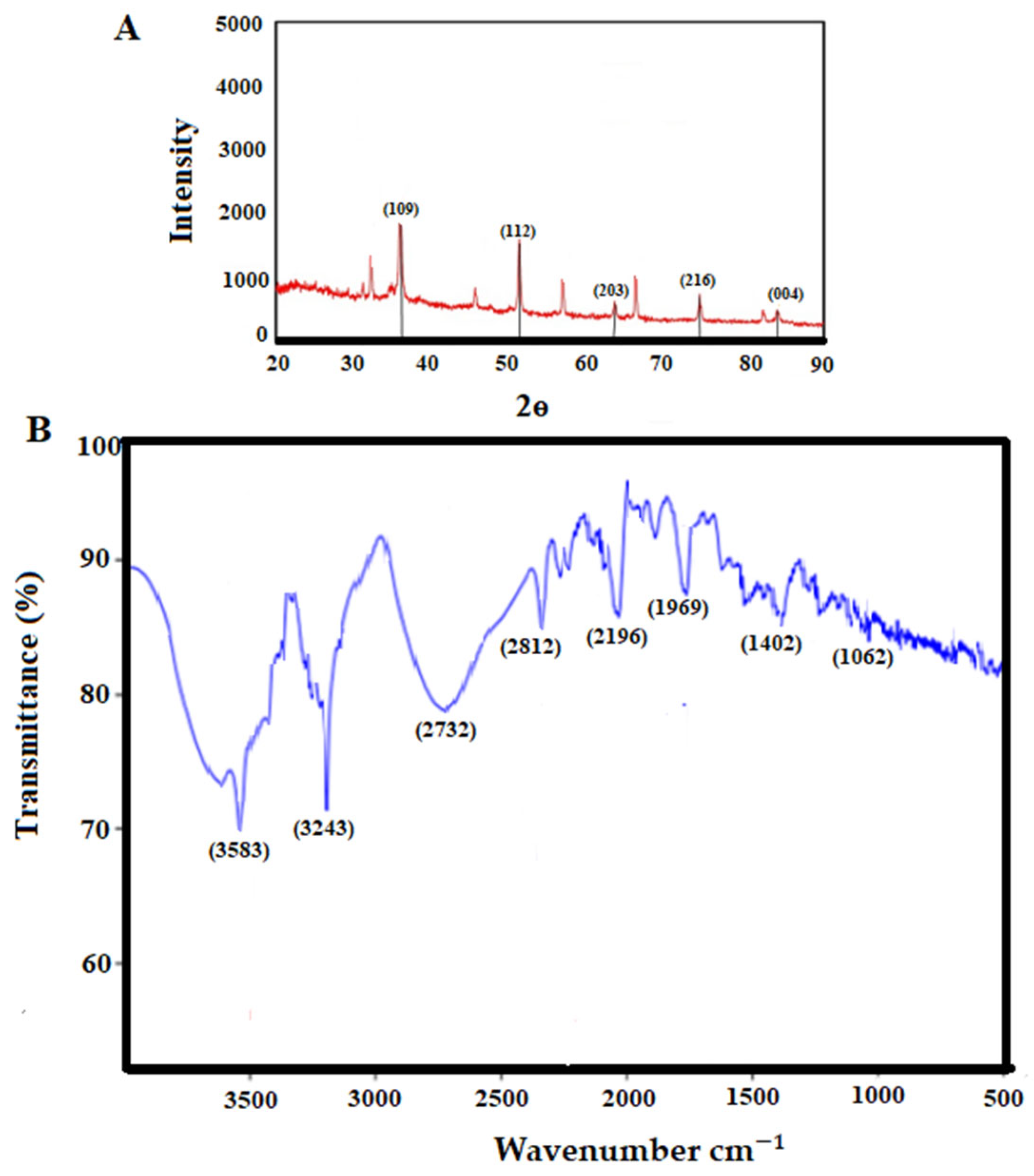
Figure 2. The analysis of X-ray diffraction (A) and Fourier transform infrared spectroscopy of the obtained copper nanoparticles (B).
FTIR spectroscopy is used to assess the function of reactants in the production of nanoparticles. In fact, the FTIR spectrum reveals the functional groups of the coating and stabilizing chemicals involved in nanoparticle creation. These compounds significantly improve the biological performance of nanoparticles [24]. The bands at 3583, 3243, 2312, 2196, 1769, 1402, and 1062 cm−1 are related to the reaction of the plant extract with copper ions, O-H stretching of alcohol and phenol, C-H stretching of the aliphatic group, C=O stretching of ester carbonyl, C=C stretching of the aromatic ring, and C-O stretching of ester, respectively (Figure 2B). This demonstrates that the biomolecules of the extract play a role both as a reducer and as a coating for copper nanoparticles, so they can protect the CNP against oxidation and transformation into copper oxide.
Figure 3 and Figure 4 show the inhibitory effects of CNP on T. gondii tachyzoite forms and THP-1 cells. CNP dose and time dependence diminished the viability of tachyzoites and THP-1 cells compared to normal saline. The IC50 value of CNP and PYM against tachyzoites was 37.2 and 25.7 µg/mL, respectively. The CC50 value of CNP and PYM on THP-1 cells was 491.4 μg/mL and 269.5 μg/mL, respectively. Following the exposure of THP-1 cells to CNP at 12.5, 25, 50, and 100 µg/mL, the rate of infectivity was evidently (p < 0.05) reduced by 68.8, 49.8, 30.2, and 11.9%, respectively; this exposure significantly reduced the number of intracellular parasites in THP-1 cells. In a study, Malekifard et al. (2020) showed the potent in vitro inhibitory effects of CNP 0.6 mg/mL against Giardia deodenalis cyst [25]. Albalawi et al. (2021) also reported that green synthesized CuNPs significantly inhibited the growth rate of Leishmania major amastigotes in a dose-dependent manner with an IC50 value of 116.8 μg/mL [26]. Another study conducted by Saad et al. (2015) demonstrated that CNP significantly reduced the viability of Entamoeba histolytica cysts and Cryptosporidium parvum oocysts with LC50-3 h of 0.13 and 0.72 mg/l, respectively [27]. These variations in the obtained results are associated with the type of tested parasites, the synthesis method of nanoparticles, and the type of assessment test [26]. Considering the antimicrobial mechanisms of action of CNP, previous studies reported that these nanoparticles displayed antimicrobial effects by provoking reactive oxygen species, disrupting cell walls, increasing cell membrane permeability, disrupting protein and DNA synthesis, restricting cell division by hindering the creation of the cell-septum-forming ring, as well as provoking the caspases activity and subsequently triggering programmed cell death [27]. The reported antimicrobial effect of CNP is not merely due to their release of metal ions but can also be attributed to their morphology, mainly their small size and high surface area to volume ratio, which allows them to interact closely with the microbial membranes of each bacterium [28].
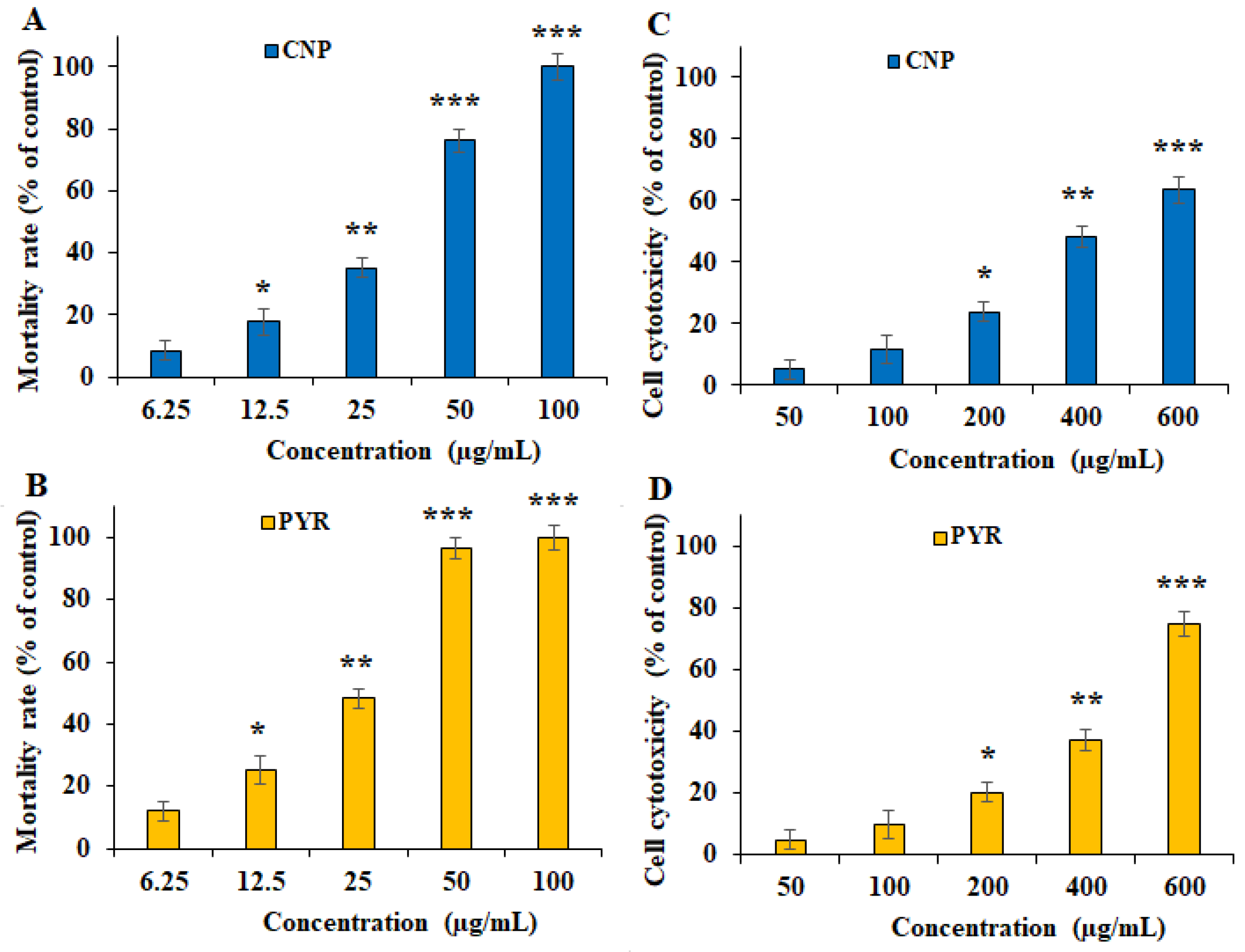
Figure 3. In vitro inhibitory effects of green synthesized copper (CNP) against Toxoplasma gondii tachyzoites (A) and THP-1 cells (C) compared to the pyrimethamine against T. gondii tachyzoites (B) and THP-1 cells (D). * p < 0.01, ** p < 0.01, and *** p < 0.001 compared to the control.
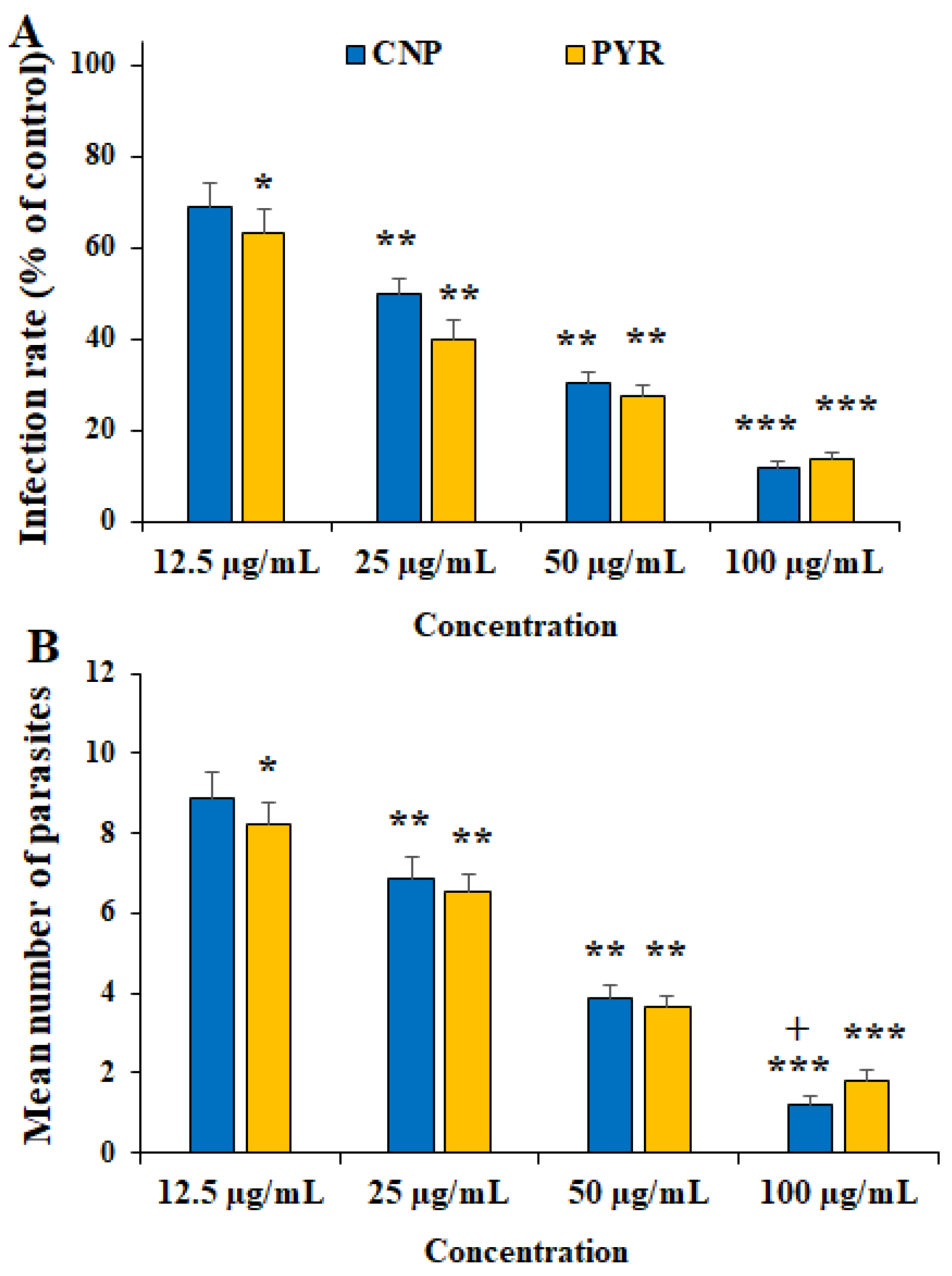
Figure 4. Effect of copper nanoparticles (CNP) and pyrimethamine (PYM) on the infectivity rate (A) and parasites inside THP-1 cells (B). * p < 0.05, ** p < 0.01, and *** p < 0.001 compared to the control (normal saline). + p < 0.05 compared to the PYM.
Figure 5 shows the effects of CLN alone and in combination with PYM on the number and size of the tissue cysts in T. gondii-infected mice. CNP at the doses of 5 and 10 mg/kg principally along with PYM noticeably (p < 0.05) reduced the number and size of the T. gondii cysts in the infected mice. With respect to the effects of green synthesized CNP against pathogenic parasites, Ezzatkhah et al. (2021) showed that green synthesized CNP using Capparis spinosa extract at 75 mg/mL along with albendazole killed the Echinococcus granulosus protoscoleces by provoking the caspase activity of this parasite [29]. In addition, Albalawi et al. revealed that green synthesized CNP alone and along with glucantime obviously inhibited and controlled the replication of Leishmania major promastigote and amastigote stages, whereas predominantly recovered the cutaneous Leishmania lesions in BALB/c mice [26]. Consistent with the study conducted by Albalawi et al. (2021), the treatment of mice infected with T. gondii Tehran strain by green synthesized CNP at doses of 2 and 4 mg/kg for two weeks markedly controlled the T. gondii infection in mice by reducing the number and size of tissue cysts [29]. Our results, in parallel with other antiparasitic studies of CNP, showed that CNP had potent in vivo antiparasitic efficacy against some pathogenic parasite strains; however, these differences in the obtained results are due to some factors, such as the type of parasite, the synthesis method of nanoparticles, and the doses used [26,30].
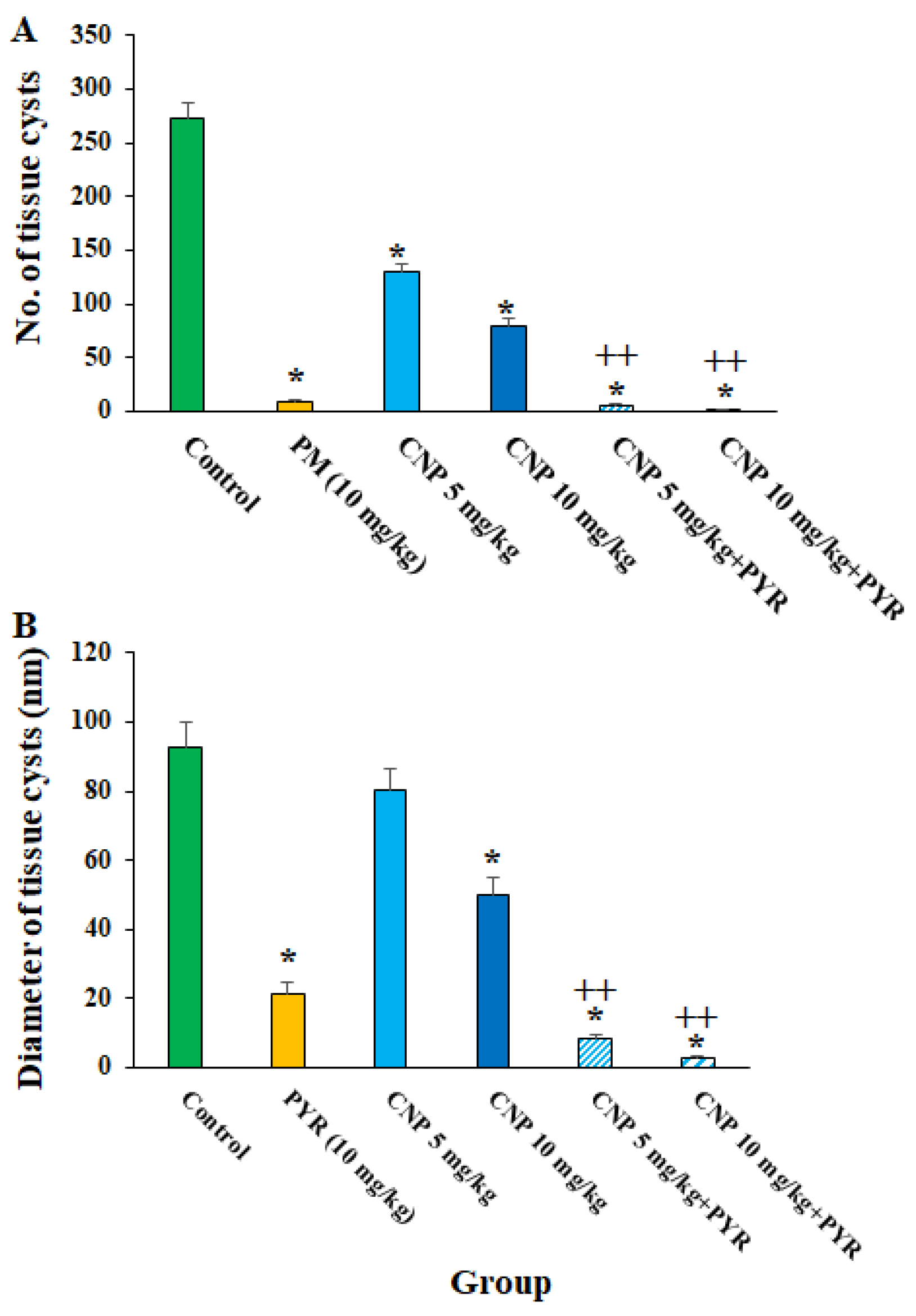
Figure 5. The number (A) and size (B) of the T. gondii cysts after treatment with copper nanoparticles (CNP) at the doses of 5 and 10 mg/kg alone in combination with pyrimethamine (5 mg/kg) for two weeks. * p < 0.001 compared to the control (normal saline). ++ p < 0.01 compared to the PYM.
Nitric oxide is the main metabolite released by various immune cells after being triggered by cytokines, which play a key function in the protection against several parasites, especially intracellular ones [31]. Accordingly, the production of cell-mediated immunity and NO triggering is well-known as a promising strategy for the design and discovery of new agents for the prevention and treatment of parasitic infections [31]. NO was produced by IFN-γ, a crucial cytokine that controls toxoplasmosis both in vitro and in vivo [32]. Our results showed that after the treatment of THP-1 cells with CNP at 1/3 IC50, ½ IC50, and IC50, the NO level was 4.13 ± 0.33, 8.5 ± 1.15, and 12.3 ± 1.55 nM (p < 0.05), respectively; this value for the cells treated with normal saline and IFN-γ + LPS was 3.24 ± 0.16 and 29.7 ± 4.15 nM, respectively. As depicted in Figure 6, following the treatment of T. gondii-infected mice with CNP, a considerable upregulation of iNOS and IFN-γ genes was observed (p < 0.001), predominantly at ½ IC50 and IC50, compared to the mice treated with normal saline (Figure 6). Consistent with our results, Albalawi et al. (2021) showed that the treatment of mice infected with T. gondii Tehran strain by green synthesized CNP at doses of 2 and 4 mg/kg for two weeks significantly controlled the TGI by increasing the expression of cellular immunity cytokines IFN-γ, IL-12, and iNO [26]. Currently, targeted host-directed immunotherapy aimed at activating or suppressing specific elements of the immune system is considered for the treatment of toxoplasmosis [33]. Considering the effect of CNP on strengthening the immune system, previous studies showed that CNP displayed their strengthening effect on the immune system via the maturation of dendritic cells and by provoking the release of cytokines IL-12 and T-cell derived cytokines IFN-γ, IL-4, IL-6, TNF-α, and IL-1β [34]. This suggests that CNP can control and prevent T. gondii infection in mice by activating cellular-mediated immunity (the central immunity factor against T. gondii).
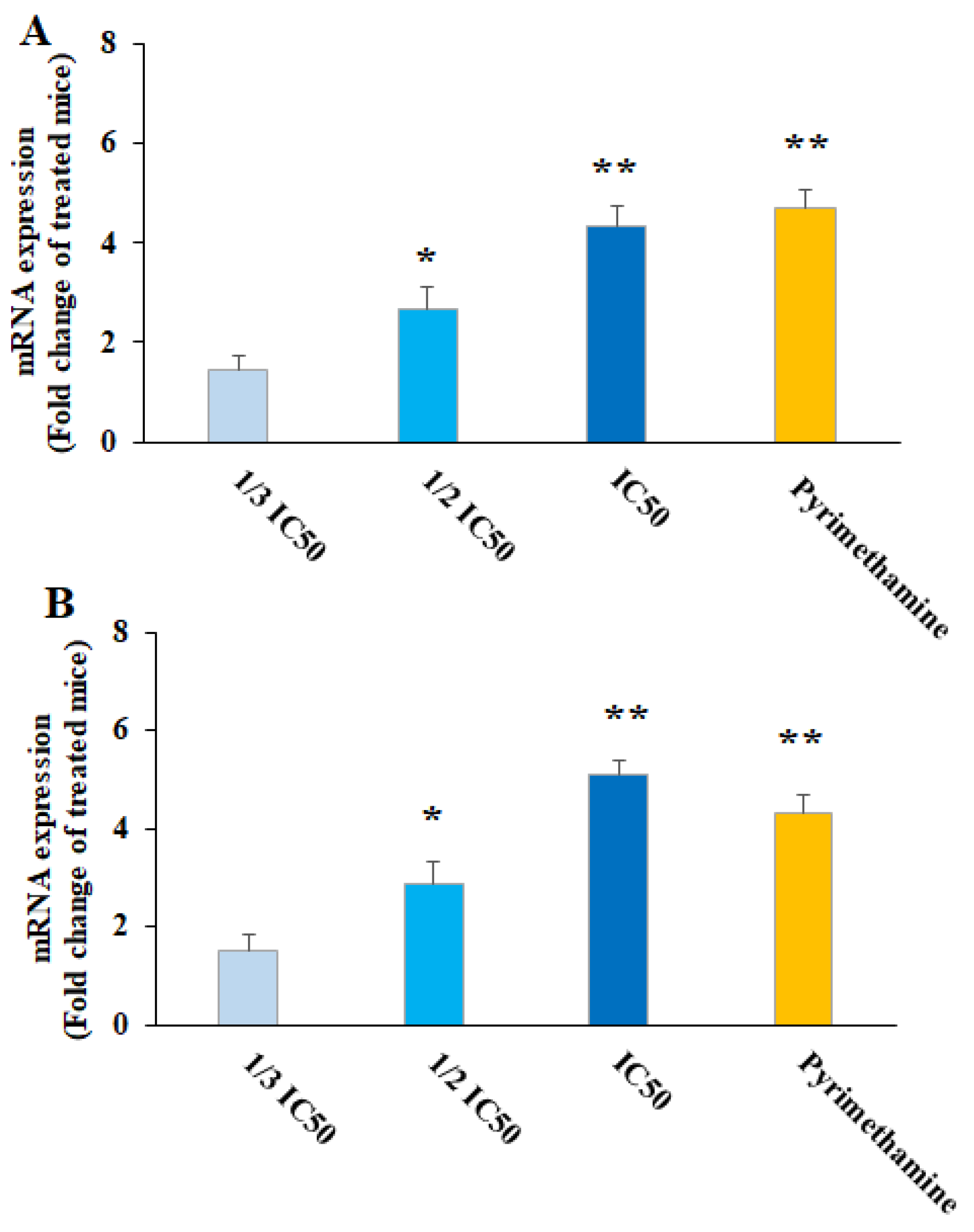
Figure 6. Effect of copper nanoparticles (CNP) and pyrimethamine (PYM) on iNOS (A) and IFN-γ (B) gene expression in THP-A cells. * p < 0.01 and ** p < 0.001 compared with control normal saline group.
Oxidative stress (OS) plays a crucial role in the pathogenesis of TGI in the host [35]. Once TGI starts, tissue damage occurs by provoking LPO production and accordingly spreading the free radicals [36]. We found that CNP at 5 and 10 mg/kg, mostly along with PYM, apparently (p < 0.05) diminished the oxidative stress in the liver of TGI mice; however, these treatments caused a significant elevation in the antioxidant factors of SOD and GPx (Figure 7), thereby demonstrating that CNP controlled the TGI in mice by decreasing the OS and increasing the antioxidant system. Similarly, Tavakoli et al. (2023) demonstrated the promising anti-inflammatory and antioxidant effects of green synthesized CNP by Artemisia annua extract in mice with second-degree burns by increasing the activity of SOD, CAT, and GPX enzymes as well as significantly reducing the MDA level [37]. It has been proven that the enzymatic antioxidant defense is one of the mechanisms that protect the host cells against an excess of free radicals due to parasitic infections such as T. gondii [38]. Since the antioxidant effects of CNP have been proven to control oxidative stress both in the present study and in previous studies, it can be suggested that these nanoparticles are able to control T. gondii infection in mice by inhibiting oxidative stress.
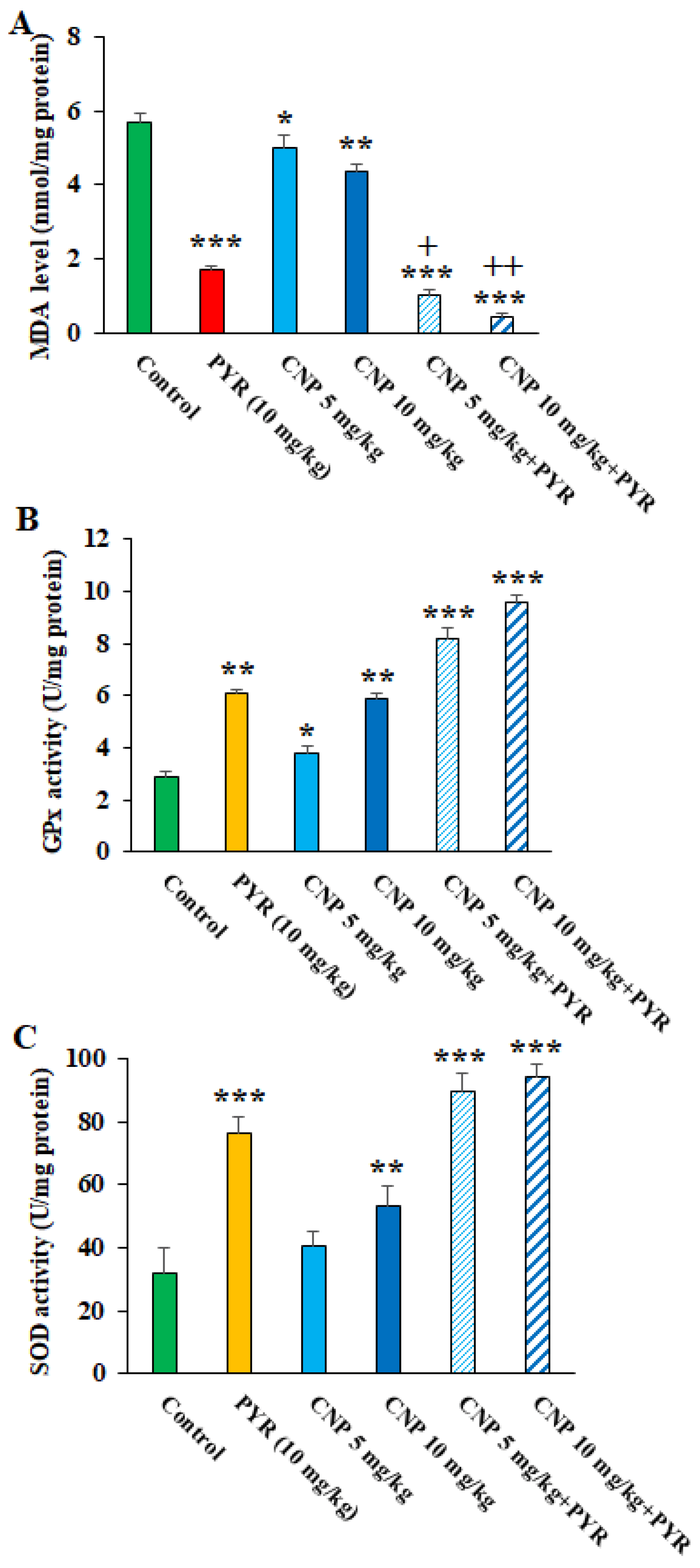
Figure 7. Effects of copper nanoparticles (CNP) treatment on the malondialdehyde (A), glutathione peroxidase (B), and superoxide dismutase enzyme activity (C) in infected mice. * p < 0.05, ** p < 0.01, and *** p < 0.001 compared to the control (normal saline). + p < 0.05 and ++ p < 0.01 compared to the pyrimethamine (PYM).
Based on the molecular tests, although TGI resulted in an increase in the expression of the BAG1 gene, following the treatment of TGI mice with CNP at 5 and 10 mg/kg alone and along with PYM, the expression of the BAG1 gene (p < 0.05) was evidently decreased (Figure 8). Previously, it was shown that disorder and reduction in the T. gondii bradyzoite-specific gene BAG1 results in significant reductions in in vivo cyst formation and bradyzoite differentiation [36]. Therefore, it can be proposed that CNP is probably able to control Toxoplasma infection in mice by reducing BAG1 gene expression and subsequently reducing the formation of tissue cysts.
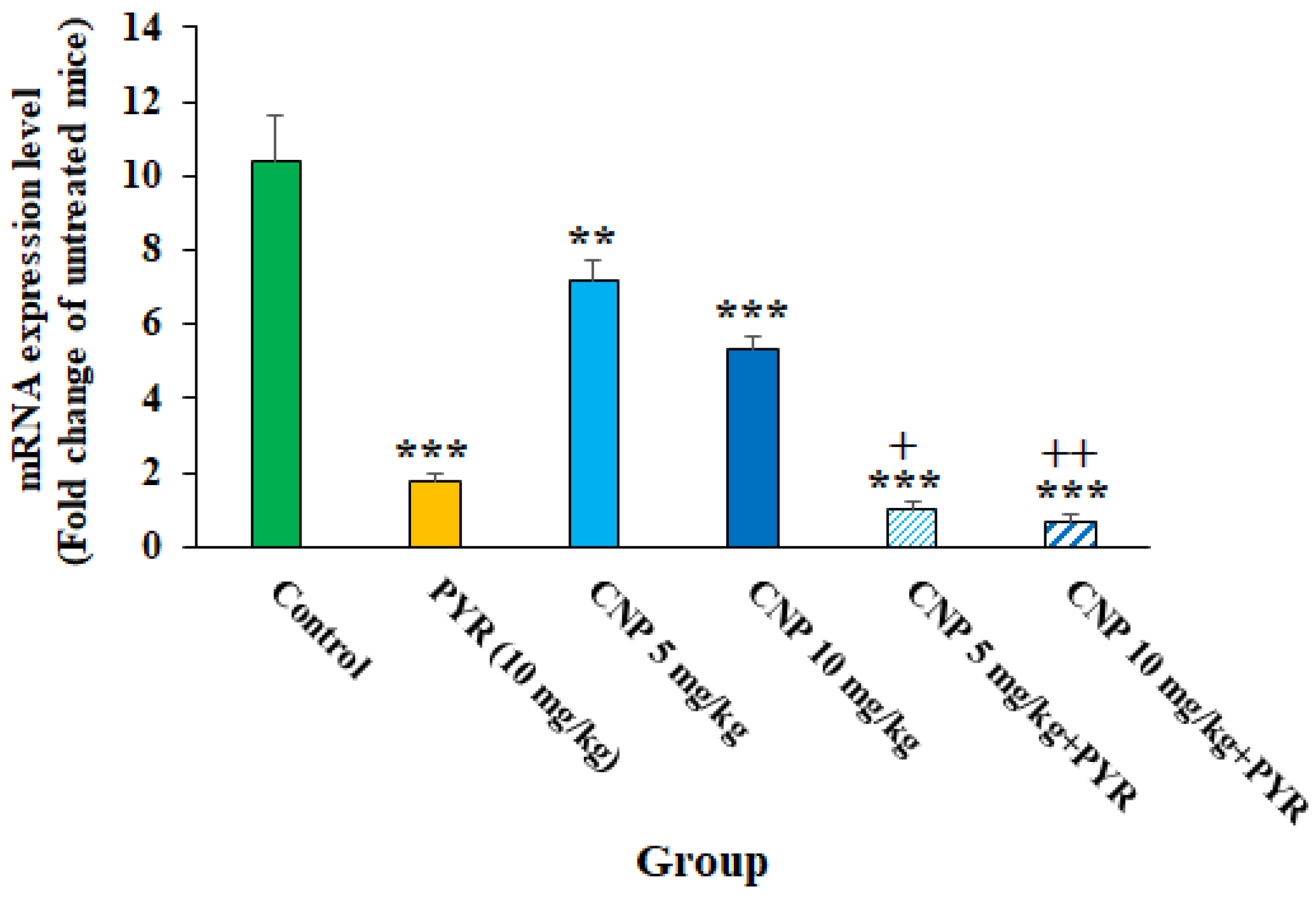
Figure 8. The effects of copper nanoparticles (CNP) alone and in combination with pyrimethamine (PYM) on the expression of the BAG1 gene in infected mice; ** p < 0.01 and *** p < 0.001 compared to the control (normal saline); + p < 0.05 compared to PYM; ++ p < 0.01 compared to PYM.
Currently, it has been proven that some factors such as the different methods of synthesis, which lead to the size, shape, and other different physical and chemical properties of the NP, can affect the toxicity of the synthesized nanoparticles [39,40]. Therefore, toxicity and optimal doses for administration should be evaluated in animal models. Regarding the toxicity effects of CNP on the markers of liver and kidney function in healthy mice, the biochemical tests demonstrated that although in some cases an increase was observed, these differences were not significant compared to the mice that received normal saline (Figure 9). Similarly, Sulaiman et al. (2018) showed that the oral administration of green synthesized Cu NPs to male Swiss albino mice had no significant toxicity in the liver, kidney, spleen, and body weight up to 400 mg/kg [41]. Accordingly, we can suggest that the oral administration of these green synthesized nanoparticles at doses of 5 and 10 mg/kg for 14 days had no toxicity on the function liver and kidney of the tested mice.
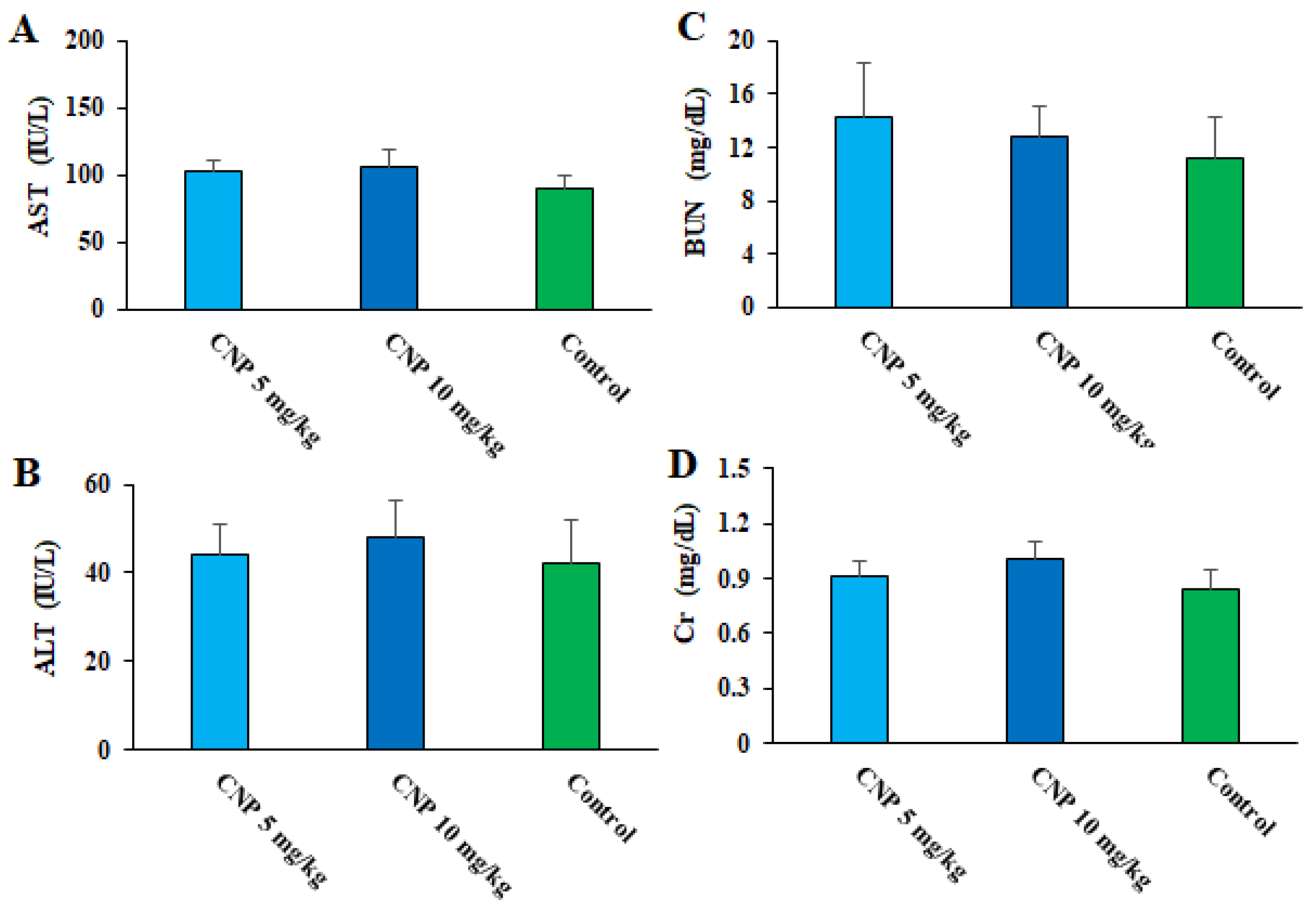
Figure 9. Effects of copper nanoparticles (CNP) on liver functional factors of aspartate transaminase (A) and alanine transaminase (B), as well as some kidney functional factors such as blood urea nitrogen (C) and creatinine (D) in healthy mice.
3. Materials and Methods
3.1. Green Synthesis of CNP
3.2. Parasite
T. gondii (ME49) was obtained from Shaqra University, Saudi Arabia, and was passaged and kept via intraperitoneal infusion in BALB/c mice.
3.3. Cell Culture
The human macrophage cell lines (THP-1) were cultured in a RPMI-1640 medium enriched with fetal bovine serum (10%) and pen/strep antibiotics (100 units/mL) at 37 °C with 5% CO2.
3.4. In Vitro Anti-Toxoplasma Effects of CNP
3.5. In Vivo Anti-Toxoplasma Effects
3.6. Safety of the Green Synthesized CNP
Thirty healthy mice were allocated into three groups (with 10 mice per group): (i) receiving normal saline for 14 days; (ii) receiving CNP daily at 5 mg/kg for 14 days; and (iii) receiving CNP daily at 10 mg/kg for 14 days. Twenty-four hours post-treatment, the animals were anesthetized utilizing a combination of ketamine and xylazine at a ratio of 100:10 mg/kg. After collecting the blood samples from the hearts of the tested mice and obtaining the serum of specimens, they were tested using commercial kits of Roche, Germany, to diagnose kidney factors of creatinine and blood urea nitrogen as well as liver function markers, such as ALT and AST [50].
3.7. Statistical Analysis
All trials were performed in triplicate to increase the reliability of the results. The acquired data were analyzed using SPSS software version 26.0, whereas ANOVA and a t-test were applied to measure the investigated groups. p < 0.05 was finally measured as a significant difference.
4. Conclusions
Our experimental study reported the beneficial effects of CNP principally along with existing chemical drugs against latent TGI in mice, whereas the possible action mechanisms of CNP include controlling oxidative stress, refining antioxidant enzymes, and increasing the production of pro-inflammatory cytokines with no toxicity to the function of vital organs. Nonetheless, additional trials to confirm these results, as well as to clarify the accurate mechanisms and their toxicity, are mandatory.
References
- Saadatnia, G.; Golkar, M. A review on human toxoplasmosis. Scand. J. Infect. Dis. 2012, 44, 805–814. [Google Scholar] [CrossRef]
- Molan, A.; Nosaka, K.; Hunter, M.; Wang, W. Global status of Toxoplasma gondii infection: Systematic review and prevalence snapshots. Trop. Biomed. 2019, 36, 898–925. [Google Scholar]
- Martinez, V.O.; Lima, F.W.d.M.; de Carvalho, C.F.; Menezes-Filho, J.A. Toxoplasma gondii infection and behavioral outcomes in humans: A systematic review. Parasitol. Res. 2018, 117, 3059–3065. [Google Scholar] [CrossRef] [PubMed]
- Hampton, M.M. Congenital Toxoplasmosis: A Review. Neonatal Netw. 2015, 34, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Tiao, N.; Gebreyes, W.A.; Jones, J.L. A review of toxoplasmosis in humans and animals in Ethiopia. Epidemiol. Infect. 2012, 140, 1935–1938. [Google Scholar] [CrossRef] [PubMed]
- McCabe, R.E. Antitoxoplasma chemotherapy. In Toxoplasmosis: A Comprehensive Clinical Guide; McCabe RE: Stockholm, Sweden, 2001; pp. 319–359. [Google Scholar]
- Montazeri, M.; Mehrzadi, S.; Sharif, M.; Sarvi, S.; Tanzifi, A.; Aghayan, S.A.; Daryani, A. Drug Resistance in Toxoplasma gondii. Front. Microbiol. 2018, 9, 2587. [Google Scholar] [CrossRef]
- Jones, J.L.; Lopez, A.; Wilson, M.; Schulkin, J.; Gibbs, R. Congenital Toxoplasmosis: A Review. Obstet. Gynecol. Surv. 2001, 56, 296–305. [Google Scholar] [CrossRef]
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of Toxoplasmosis: Historical Perspective, Animal Models, and Current Clinical Practice. Clin. Microbiol. Rev. 2018, 31, e00057-17. [Google Scholar] [CrossRef]
- Marra, C.M. Central nervous system infection with Toxoplasma gondii. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 152, pp. 117–122. [Google Scholar]
- Formoso, P.; Muzzalupo, R.; Tavano, L.; De Filpo, G.; Nicoletta, F.P. Nanotechnology for the Environment and Medicine. Mini Rev. Med. Chem. 2016, 16, 668–675. [Google Scholar] [CrossRef]
- Mahmoudvand, H.; Mohebali, M.; Sharifi, I.; Keshavarz, H.; Hajjaran, H.; Akhoundi, B.; Jahanbakhsh, S.; Zarean, M.; Javadi, A. Epidemiological aspects of visceral leishmaniasis in Baft district, Kerman Province, Southeast of Iran. Iran. J. Parasitol. 2011, 6, 1–11. [Google Scholar]
- Din, M.I.; Rehan, R. Synthesis, Characterization, and Applications of Copper Nanoparticles. Anal. Lett. 2016, 50, 50–62. [Google Scholar] [CrossRef]
- Khodashenas, B.; Ghorbani, H.R. Synthesis of copper nanoparticles: An overview of the various methods. Korean J. Chem. Eng. 2014, 31, 1105–1109. [Google Scholar] [CrossRef]
- Rafique, M.; Shaikh, A.J.; Rasheed, R.; Tahir, M.B.; Bakhat, H.F.; Rafique, M.S.; Rabbani, F. A review on synthesis, characterization and applications of copper nanoparticles using green method. Nano 2017, 12, 1750043. [Google Scholar] [CrossRef]
- Graham, S.A.; Turkington, R. Population dynamics response of Lupinus arcticus to fertilization, clipping, and neighbour removal in the understory of the boreal forest. Can. J. Bot. 2000, 78, 753–758. [Google Scholar] [CrossRef]
- Ingham, J.L.; Tahara, S.; Harborne, J.B. Fungitoxic Isoflavones from Lupinus albus and other Lupinus Species. Z. Naturforsch. C 1983, 38, 194–200. [Google Scholar] [CrossRef]
- Cheraghipour, K.; Masoori, L.; Ezzatkhah, F.; Salimikia, I.; Amiri, S.; Makenali, A.S.; Taherpour, F.; Mahmoudvand, H. Effect of chitosan on Toxoplasma gondii infection: A systematic review. Parasite Epidemiol. Control. 2020, 11, e00189. [Google Scholar] [CrossRef]
- Saadatmand, M.; Al-Awsi, G.R.L.; Alanazi, A.D.; Sepahvand, A.; Shakibaie, M.; Shojaee, S.; Mohammadi, R.; Mahmoudvand, H. Green synthesis of zinc nanoparticles using Lavandula angustifolia Vera. Extract by microwave method and its prophylactic effects on Toxoplasma gondii infection. Saudi J. Biol. Sci. 2021, 28, 6454–6460. [Google Scholar] [CrossRef]
- Pestryakov, A.; Petranovskii, V.; Kryazhov, A.; Ozhereliev, O.; Pfänder, N.; Knop-Gericke, A. Study of copper nanoparticles formation on supports of different nature by UV-Vis diffuse reflectance spectroscopy. Chem. Phys. Lett. 2004, 385, 173–176. [Google Scholar] [CrossRef]
- Khodaie, M.; Ghasemi, N. Green synthesis and characterization of copper nanoparticles using Eryngium campestre leaf extract. Bulg. Chem. Comm. 2018, 50, 244–250. [Google Scholar]
- Xu, J.; Song, M.; Fang, Z.; Zheng, L.; Huang, X.; Liu, K. Applications and challenges of ultra-small particle size nanoparticles in tumor therapy. J. Control. Release 2022, 353, 699–712. [Google Scholar] [CrossRef]
- Holder, C.F.; Schaak, R.E. Tutorial on Powder X-ray Diffraction for Characterizing Nanoscale Materials. ACS Nano 2019, 13, 7359–7365. [Google Scholar] [CrossRef] [PubMed]
- Berthomieu, C.; Hienerwadel, R. Fourier transform infrared (FTIR) spectroscopy. Photosynth. Res. 2009, 101, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Malekifard, F.; Tavassoli, M.; Vaziri, K. In Vitro Assessment Antiparasitic Effect of Selenium and Copper Nanoparticles on Giardia deodenalis Cyst. Iran. J. Parasitol. 2020, 15, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Albalawi, A.E.; Abdel-Shafy, S.; Khalaf, A.K.; Alanazi, A.D.; Baharvand, P.; Ebrahimi, K.; Mahmoudvand, H. Therapeutic Potential of Green Synthesized Copper Nanoparticles Alone or Combined with Meglumine Antimoniate (Glucantime®) in Cutaneous Leishmaniasis. Nanomaterials 2021, 11, 891. [Google Scholar] [CrossRef] [PubMed]
- Saad, A.H.; Soliman, M.I.; Azzam, A.M.; Mostafa, A.B. Antiparasitic activity of silver and copper oxide nanoparticles against Entamoeba histolytica and Cryptosporidium parvum cysts. J. Egypt. Soc. Parasitol. 2015, 45, 593–602. [Google Scholar]
- Betancourt-Galindo, R.; Reyes-Rodriguez, P.Y.; Puente-Urbina, B.A.; Avila-Orta, C.A.; Rodríguez-Fernández, O.S.; Cadenas-Pliego, G.; Lira-Saldivar, R.H.; García-Cerda, L.A. Synthesis of Copper Nanoparticles by Thermal Decomposition and Their Antimicrobial Properties. J. Nanomater. 2014, 2014, 980545. [Google Scholar] [CrossRef]
- Albalawi, A.E.; Alanazi, A.D.; Alyousif, M.S.; Sepahvand, A.; Ebrahimi, K.; Niazi, M.; Mahmoudvand, H. The High Potency of Green Synthesized Copper Nanoparticles to Prevent the Toxoplasma gondii Infection in Mice. Acta Parasitol. 2021, 66, 1472–1479. [Google Scholar] [CrossRef]
- Ma, X.; Zhou, S.; Xu, X.; Du, Q. Copper-containing nanoparticles: Mechanism of antimicrobial effect and application in dentistry—A narrative review. Front. Surg. 2022, 9, 905892. [Google Scholar] [CrossRef]
- James, S.L. Role of nitric oxide in parasitic infections. Microbiol. Rev. 1995, 59, 533–547. [Google Scholar] [CrossRef]
- Gazzinelli, R.; Brezin, A.; Li, Q.; Nussenblatt, R.; Chan, C. Toxoplasma gondii: Acquired Ocular Toxoplasmosis in the Murine Model, Protective Role of TNF-α and IFN-γ. Exp. Parasitol. 1994, 78, 217–229. [Google Scholar] [CrossRef]
- Gazzinelli, R.; Denkers, E.Y.; Sher, A. Host resistance to Toxoplasma gondii: Model for studying the selective induction of cell-mediated immunity by intracellular parasites. Infect. Agents Dis. 1993, 2, 139–149. [Google Scholar] [PubMed]
- Tulinska, J.; Mikusova, M.L.; Liskova, A.; Busova, M.; Masanova, V.; Uhnakova, I.; Rollerova, E.; Alacova, R.; Krivosikova, Z.; Wsolova, L.; et al. Copper Oxide Nanoparticles Stimulate the Immune Response and Decrease Antioxidant Defense in Mice after Six-Week Inhalation. Front. Immunol. 2022, 13, 874253. [Google Scholar] [CrossRef] [PubMed]
- Nazarlu, Z.H.A.; Matini, M.; Bahmanzadeh, M.; Foroughi-Parvar, F. Toxoplasma gondii: A Possible Inducer of Oxidative Stress in Reproductive System of Male Rats. Iran. J. Parasitol. 2020, 15, 521–529. [Google Scholar] [CrossRef]
- Tavakoli, F.; Karimi Babaahmadi, B.; Javdani, M.; Kaboutari, J. Antioxidant effect of copper nanoparticles coated with Artemisia annua plant extract in the 2nd degree burns in mice. J. Comp. Pathobiol. 2023, 20, 4065–4074. [Google Scholar]
- Szewczyk-Golec, K.; Pawłowska, M.; Wesołowski, R.; Wróblewski, M.; Mila-Kierzenkowska, C. Oxidative Stress as a Possible Target in the Treatment of Toxoplasmosis: Perspectives and Ambiguities. Int. J. Mol. Sci. 2021, 22, 5705. [Google Scholar] [CrossRef]
- Louis, M.W.; Weiss, L.M.; Kim, K. The development and biology of bradyzoites of Toxoplasma gondii. Front. Biosci. 2000, 5, 391–405. [Google Scholar] [CrossRef]
- Zoroddu, M.A.; Medici, S.; Ledda, A.; Nurchi, V.M.; Lachowicz, J.I.; Peana, M. Toxicity of nanoparticles. Curr. Med. Chem. 2014, 21, 3837–3853. [Google Scholar] [CrossRef]
- Sulaiman, G.M.; Tawfeeq, A.T.; Jaaffer, M.D. Biogenic synthesis of copper oxide nanoparticles using Olea europaea leaf extract and evaluation of their toxicity activities: An in vivo and in vitro study. Biotechnol. Prog. 2018, 34, 218–230. [Google Scholar] [CrossRef]
- Mahmoudvand, H.; Sepahvand, P.; Jahanbakhsh, S.; Azadpour, M. Evaluation of the antileishmanial and cytotoxic effects of various extracts of garlic (Allium sativum) on Leishmania tropica. J. Parasit. Dis. 2016, 40, 423–426. [Google Scholar] [CrossRef]
- Jahanbakhsh, S.; Azadpour, M.; Tavakoli Kareshk, A.; Keyhani, A.; Mahmoudvand, H. Zataria multiflora Bioss: Lethal effects of methanolic extract against protoscoleces of Echinococcus granulosus. J. Parasit. Dis. 2016, 40, 1289–1292. [Google Scholar] [CrossRef]
- Ezzatkhah, F.; Khalaf, A.K.; Mahmoudvand, H. Copper nanoparticles: Biosynthesis, characterization, and protoscolicidal effects alone and combined with albendazole against hydatid cyst protoscoleces. Biomed. Pharmacother. 2021, 136, 111257. [Google Scholar] [CrossRef] [PubMed]
- Ezzatkhah, F.; Mahmoudvand, H.; Raziani, Y. The role of Curcuma longa essential oil in controlling acute toxoplasmosis by improving the immune system and reducing inflammation and oxidative stress. Front. Cell. Infect. Microbiol. 2023, 13, 1161133. [Google Scholar] [CrossRef] [PubMed]
- Yadegari, J.G.; Khalaf, A.K.; Ezzatkhah, F.; Shakibaie, M.; Mohammadi, H.R.; Mahmoudvand, H. Antileishmanial, cellular mechanisms, and cytotoxic effects of green synthesized zinc nanoparticles alone and in combined with glucantime against Leishmania major infection. Biomed. Pharmacother. 2023, 164, 114984. [Google Scholar] [CrossRef]
- De Oliveira, T.C.; Silva, D.A.; Rostkowska, C.; Béla, S.R.; Ferro, E.A.; Magalhães, P.M.; Mineo, J.R. Toxoplasma gondii: Effects of Artemisia annua L. on susceptibility to infection in experimental models in vitro and in vivo. Exp. Parasitol. 2009, 122, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudvand, H.; Yadegari, J.G.; Khalaf, A.K.; Hashemi, M.J.; Dastyarhaghighi, S.; Salimikia, I. Chemical composition, antileishmanial, and cytotoxic effects Ferula macrecolea essential oil against Leishmania tropica. Parasite Epidemiol. Control 2022, 19, e00270. [Google Scholar] [CrossRef]
- Shaapan, R.M.; Al-Abodi, H.R.; Alanazi, A.D.; Abdel-Shafy, S.; Rashidipour, M.; Shater, A.F.; Mahmoudvand, H. Myrtus communis Essential Oil; Anti-Parasitic Effects and Induction of the Innate Immune System in Mice with Toxoplasma gondii Infection. Molecules 2021, 26, 819. [Google Scholar] [CrossRef]
- Keyhani, A.; Ziaali, N.; Shakibaie, M.; Kareshk, A.T.; Shojaee, S.; Asadi-Shekaari, M.; Sepahvand, M.; Mahmoudvand, H. Biogenic selenium nanoparticles target chronic toxoplasmosis with minimal cytotoxicity in a mouse model. J. Med. Microbiol. 2020, 69, 104–110. [Google Scholar] [CrossRef]
- Mahmoudvand, H.; Pakravanan, M.; Aflatoonian, M.R.; Khalaf, A.K.; Niazi, M.; Mirbadie, S.R.; Tavakoli Kareshk, A.; Khatami, M. Efficacy and safety of Curcuma longa essential oil to inactivate hydatid cyst protoscoleces. BMC Complement. Altern. Med. 2019, 19, 187. [Google Scholar] [CrossRef]
