


1. Introduction
Adult-type diffuse gliomas represent one of the most common types of central nervous system tumors and occur in 5–6 cases per 100,000. Currently, the first-line treatment for grade III–IV gliomas is surgical resection followed by radiotherapy and chemotherapy temozolomide, but this does not significantly increase the life expectancy of patients [1].
Therapy with oncolytic viruses is an actively developing approach to cancer treatment, allowing for the selective targeting and lysis of tumor cells [2]. In addition to direct oncolysis, virotherapy induces an antitumor immune response. Tumor-associated antigens are released during cell lysis, leading to their recognition by the immune system and the recruitment of activated immune cells [3]. At present, various oncolytic viruses are undergoing preclinical and clinical trials as therapies for glioblastoma. Moreover, the recent approval of the oncolytic herpes virus G47∆ as a pioneering glioblastoma treatment in Japan emphasizes the expediency of further research [4].
Underlying the development of oncolytic approaches, the vaccinia virus (VACV) has the principal advantage of a natural tropism for tumors of various histogenesis, as it does not require specific receptors to penetrate the cell [5]. A main strategy for enhancing the oncolytic effect of VACV involves inserting transgenes into the virus genome. The introduced sequences can encode proteins that induce apoptosis or mediate recruiting host immune cells and enhance the antitumor immune response. In particular, the selective targeting of VACV to a tumor is ensured by the optimal conditions for successful virus replication that are created by the hallmarks of malignant cell transformation (e.g., disturbances in the apoptotic pathways, cell cycle dysregulation, and immune system evasion). In addition, VACV replication and dissemination depend on the epidermal growth factor receptor (EGFR) signaling pathway, which is activated in most tumors [6].
JX-594, or Pexa-Vec, is one of the most well-known and studied recombinant VACVs used as a candidate drug for the treatment of malignant neoplasms [7]. This modified virus lacks the viral thymidine kinase (tk) gene but carries human GM-CSF and β-galactosidase genes under the control of synthetic early/late and p7.5 promoters, respectively. The inactivation of the thymidine kinase gene makes virus replication dependent on high thymidine kinase activity in the cell, a characteristic feature of tumor cells [8]. GM-CSF expressed by the virus stimulates the antitumor immune response [9]. Indications for using Pexa-Vec in clinical trials include hepatocellular carcinoma, melanoma, breast cancer, and other solid tumors. The drug has demonstrated safety with various routes of administration; however, clinical approval has not yet been obtained.
Previously, we have developed VV-GMCSF-Lact, a recombinant VACV derived from the parent Lister (L-IVP) strain. VV-GMCSF-Lact is considered a promising drug for the treatment of solid tumors, including glioblastoma [10]. In this viral construct, the viral thymidine kinase (tk) and growth factor (vgf) genes are replaced with human genes encoding GM-CSF and the apoptosis-inducing protein lactaptin, respectively [10]. Lactaptin is a proteolytic fragment of human kappa-casein. It has proapoptotic activity against tumor cells, inducing apoptosis via the mitochondrial pathway and autophagy [11]. VV-GMCSF-Lact is currently undergoing clinical trials as an oncolytic treatment for breast cancer, including triple-negative (ClinicalTrials.gov Identifier: NCT05376527). In addition, we have shown that VV-GMCSF-Lact exhibits high oncolytic activity in vitro, as well as antitumor efficacy against human glioma in vivo. Its ability to penetrate the blood–brain barrier has been established both in mice with tumors and in intact mice [12]. Since cells of different glioma cultures can show different sensitivity to VV-GMCSF-Lact, it is necessary to study the mechanisms that determine the sensitivity of glioma cells to the virus in order to develop the most effective therapeutic approaches.
Here, to describe the effect of VV-GMCSF-Lact on the human cell transcriptome, we used patient-derived glioma/glioblastoma cell cultures (BR1, BR3, BR4, BR5), immortalized U87 and U343 human glioblastoma cell lines, and a patient-derived non-malignant human brain cell culture (NB). By analyzing sets of differentially expressed genes (DEGs), we determined general changes in the transcriptome and identified individual clusters of genes associated with sensitivity or resistance of glioma cells to the cytotoxic activity of VV-GMCSF-Lact.
2. Materials and Methods
2.1. Immortalized Cell Lines
Human U87 MG and U343 MG cell lines were purchased from the Russian cell culture collection (Russian Branch of the ETCS, St. Petersburg, Russia). The cells were cultivated in Minimum Essential Medium α (MEM α; Sigma-Aldrich, MS, USA) supplemented with 10% FBS (Gibco BRL Co., Gaithersburg, MD, USA), 2 mM L-glutamine (Sigma-Aldrich, MS, USA), 250 mg/mL amphotericin B, and 100 U/mL penicillin/streptomycin (Gibco BRL Co., Gaithersburg, MD, USA) at 37 °C in a humidified atmosphere containing 5% CO2.
2.2. Patient-Derived Cell Cultures
Glioma tissue samples were obtained at the Novosibirsk Research Institute of Traumatology and Orthopedics (Novosibirsk, Russia) from patients who provided informed consent. The study was approved by the Committee on the Ethics of the Novosibirsk Research Institute of Traumatology and Orthopedics (protocol no. 050/17 68 of 11 September 2017). A sample of normal brain tissue (NB) was obtained from a patient without a malignant tumor at the time of surgery. Glioma samples (BR1, BR3, BR4, and BR5) and NB were mechanically dissociated in Iscove’s modified Dulbecco’s media (IMDM, Sigma-Aldrich, MS, USA). Then, specimens were washed with 10X excess of phosphate-buffered saline (PBS) and collected with centrifugation at 300× g. Specimens were seeded in 6-well plates using IMDM medium supplemented with 10% FBS, 2 mM L-glutamine, 100 U/mL penicillin, 100 µg/mL streptomycin, and 250 mg/mL amphotericin B. They were then incubated at 37 °C in a humidified atmosphere containing 5% CO2 for cell adhesion, and the medium was changed every three to four days. After reaching 70–80% confluence, cells were harvested using Triple-Express (GIBCO, Thermo Fisher, Waltham, NY, USA) and subcultured for further experiments. In the following steps, the cells were cultured under the same conditions.
2.3. Oncolytic Virus VV-GMCSF-Lact
The recombinant VV-GMCSF-Lact was engineered from the VACV Lister strain (L-IVP) and contained deletions of the viral thymidine kinase (tk) and virus growth factor (vgf) gene fragments. In the corresponding regions, we inserted the gene of human GM-CSF (CSF2) and the gene of the proapoptotic fragment of human kappa-casein (CSN2) or lactaptin [10]. VV-GMCSF-Lact was produced in African green monkey kidney cells 4647 and purified as described in [13]. The viral titer was determined using the plaque-forming assay and expressed as a number of plaque-forming units per volume (i.e., PFU/mL) [13].
2.4. Flow Cytometry
Cells of immortalized (U87 MG and U343 MG) and patient-derived (BR1, BR3, BR4, BR5, NB) cultures grown in 6-well plates were collected and incubated with phycoerythrin (PE)-conjugated mouse anti-human CD133 mAbs (Miltenyi Biotec, Bergisch Gladbach, Germany), fluorescein isothiocyanate (FITC)-conjugated rat anti-human CD44 antibody (Invitrogen, CA, USA), Alexa Fluor 488-conjugated mouse anti-human/mouse CD15 antibody (R&D Systems, Minneapolis, MN, USA), and PE-conjugated mouse anti-human CD171 antibody (Sony Biotechnology, San Jose, CA, USA) in PBS supplemented with 0.5% fetal bovine serum and 2 mM EDTA for 30 min on ice. The assay was run on a BD FACSCantoII flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA), and the data were analyzed using the BD FACSDiva Software (BD Biosciences, Franklin Lakes, NJ, USA).
2.5. XCelligence Assay
The cell proliferation and adhesion kinetics were determined using RTCA technology (ACEA Biosciences, San Diego, CA USA). Cells of immortalized (U87 and U343) and patient-derived (BR1, BR3, BR4, BR5, NB) cultures were seeded (30,000 cells/well) in an 8-well E-plate in three technical replicates. The plate was incubated in a humidified atmosphere containing 5% CO2 at 37 °C for 72 h, and the cell index for each culture was automatically tracked during this period.
2.6. Apoptosis Detection
Cells of immortalized (U87 and U343) and patient-derived (BR1, BR3, BR4, BR5, NB) cultures were treated with VV-GMCSF-Lact at a multiplicity of infection (MOI) of 1 PFU per cell. Following 24 h incubation with the virus, cells were harvested using Triple-Express (GIBCO, Thermo Fisher, Waltham, NY, USA) and stained with annexin V-FITC and PI using the BD Pharmingen Apoptosis Detection Kit (BD Bioscience, Franklin Lakes, NJ, USA) according to the manufacturer’s protocol. Cells without any virus exposure were used as controls. The analyses were performed using the BD FACSCantoII flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA), and the data were analyzed using the BD FACSDiva Software (BD Biosciences, Franklin Lakes, NJ, USA).
2.7. Determination of the Cytotoxic Dose (CD50) of VV-GMCSF-Lact for Glioma and Normal Brain Cell Cultures
To study the cytotoxic activity of VV-GMCSF-Lact, cells of immortalized (U87 MG and U343 MG) and patient-derived (BR1, BR3, BR4, BR5, NB) cultures were plated into 96-well plates, 3000 cells per well. Then, cells were treated with VV-GMCSF-Lact at an MOI ranging from 10 to 0.0012 PFU per cell using a two-step dilution factor. After 72 h incubation at 37 °C in a humidified atmosphere containing 5% CO2, cell viability was evaluated using the Deep Blue Cell Viability™ Kit (Biolegend, CA, USA). Cell viability was determined relative to the viability of the control cells (100% ± standard deviation in three independent experiments). The cytotoxic dose (CD50, when 50% of cells die) was calculated using the Compusyn software [14].
2.8. Treatment with VV-GMCSF-Lact for Next-Generation RNA Sequencing
To obtain RNA samples for the next-generation RNA sequencing (NGS), the cells of immortalized (U87 and U343) and patient-derived (BR1, BR3, BR4, BR5, NB1) cultures were first plated into 6-well plates, with 1 million cells per well. Then, cells were treated with 1 PFU per cell of VV-GMCSF-Lact. After 12 h and 24 h incubation at 37 °C in a humidified atmosphere containing 5% CO2, cells were harvested using Triple-Express (GIBCO, Thermo Fisher, Waltham, NY, USA). Cells incubated in the same conditions but without exposure to the virus were used as the control.
2.9. RNA Isolation
To isolate total RNA, an RNA extraction kit (LRU-100-50, Biolabmix, Russia) was used according to the manufacturer’s protocol. RNA concentration was assessed using the Qubit 2 fluorometer (Thermo Fisher Scientific, USA) and the Qubit RNA HS Assay Kit (Thermo Fisher Scientific, USA). The quality of total RNA, expressed as the RNA integrity number (RIN), was determined using Bioanalyzer 2100 (Agilent, USA) with the Agilent RNA Pico 6000 Kit (Agilent, USA). A threshold RIN value greater than 8.0 was taken as the cutoff point for moving to the library preparation stage.
2.10. RNA Sequencing
Illumina cDNA libraries were produced according to a standard protocol using the NEBNext Ultra II Targeted RNA Library Preparation Kit (New England Biolabs, UK) and the NEBNext mRNA Magnetic Isolation Module (New England Biolabs, UK), as well as mass parallel sequencing on the NextSeq Illumina platform 1500 at the Institute of Fundamental Medicine and Biology of the Kazan Federal University (Kazan, Russia). For mRNA isolation, fragmentation, and priming, 1 μg of total RNA was used. The NextSeq 500/550 High Output v2.5 Kit (Illumina, USA) creating 100-nucleotide single-end reads was used. Fragment size distribution in the prepared sequencing libraries was analyzed using the Bioanalyzer 2100 instrument (Agilent, USA) with the Agilent High Sensitivity DNA Kit (Agilent, USA) and quantified using the Qubit 2.0 Fluorometer (Invitrogen, USA) with the Qubit dsDNA HS Assay Kit (Thermo Fisher Scientific, USA). Fragment sizes varied from 250 to 700 bp with a clear peak at 300 bp.
2.11. Transcriptome Analysis
Raw sequencing reads (76-nucleotide single-end reads) were subjected to Illumina adapter removal using Trimmomatic [15]. The trimmed sequencing reads were filtered with Bowtie2 [16] using a reference containing sequences of human rRNAs (RefSeq); tRNAs; snRNA; SINE-, LINE-, and DNA-repeat consensus sequences (RepBase [17]); low-complexity simple repeats; and mitochondrial DNA (NC_012920.1). The filtered reads were mapped to the human genome (GRCh38/hg38) with the addition of the VV-GMCSF-Lact genome as a single chromosome. The CSF3 and CSN3 (corresponding to lactaptin) gene sequences were excluded from the human genome by masking the corresponding regions with N. Filtered reads were mapped with STAR 2.7.1a [18] using the RefGene human genome annotation (https://hgdownload.soe.ucsc.edu/goldenPath/hg38/bigZips/genes/, accessed on 3 March 2022) modified with the VV-GMCSF-Lact genome annotation. To construct the VV-GMCSF-Lact genome annotation, we used the annotation of the parental VACV strain L-IVP (GenBank accession KP233807) and experimental data from [10]. Aligned reads were quantified using QoRTs v1.3.6 [19]. Differential gene expression analysis was performed with DESeq2 1.36.0 [20], R version 4.1.3, and Bioconductor 3.14. Following the differential gene expression analysis, lists of upregulated and downregulated genes were analyzed with Enrichr using the R interface [21].
3. Results
3.1. Cell Cultures and General NGS RNA-Seq Data
3.2. General Changes in Individual Transcript Levels, Transcription Factor Activity, Biological Processes, and Pathways Affected by VV-GMCSF-Lact Infection
To describe the overall transcriptome changes in human glioma and NB cells during infection with VV-GMCSF-Lact, we employed a cell-specific approach using DESeq2. It allowed us to compare transcriptome changes in each cell culture at 12 h and 24 h time points after infection. In cell-specific DESeq2 comparisons, we determined the number of differentially expressed transcripts (or differentially expressed genes, DEGs) for each cell culture. The highest variation in RNA patterns at 24 h was observed in the BR1 cell culture (6587 DEGs), whereas U87 and NB cells, which are relatively resistant to VV-GMCSF-Lact, demonstrated the lowest variations (1384 and 1563 DEGs, respectively) .
We constructed a heat map showing the most variable upregulated and downregulated DEGs in glioma and NB cell lines 12 and 24 h after VV-GMCSF-Lact infection; histone gene transcripts are marked with red dots (Figure 4). When compared to other cell culture parameters ( and , the number of DEGs was found to correlate well with the relative contribution of viral transcripts (R2 = 0.72, ). At the same time, we found no significant correlation between the number of DEGs and CD50 or other cell culture parameters presented in (Pearson R2 < 0.2, data not illustrated).
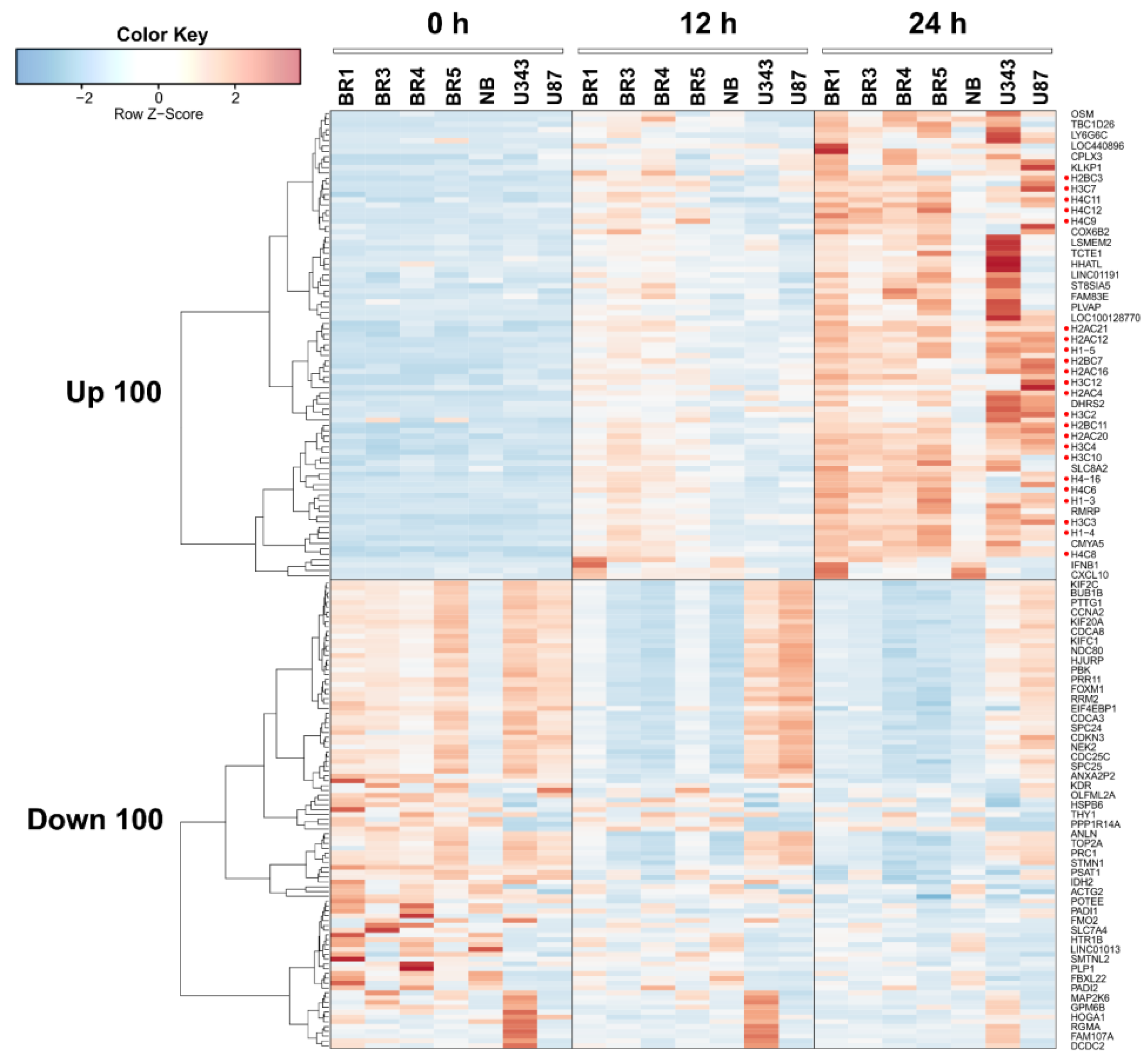
Figure 4. Heatmap of the 100 most variable upregulated and downregulated differentially expressed genes in glioma and NB cell cultures 12 h and 24 h after VV-GMCSF-Lact infection. Histone gene transcripts are marked with red dots.
Thus, the cytotoxic and proapoptotic effects of VV-GMCSF-Lact infection are not directly associated with the overall level of viral gene expression; rather, they are determined by the activation of specific host genes and/or viral genes. Yet, it is of particular interest to describe the processes triggered by the oncolytic virus that are common across different cell cultures. Therefore, here, we provide a brief description of the common genes and general processes that are activated or suppressed during VV-GMCSF-Lact infection of human glioma and NB cell cultures.
It is known that viral infection is accompanied by large-scale changes in fundamental cellular processes. Different cells in a culture, organ, or tissue react differently to the virus, activating the innate and adaptive immune response. The response to the virus infection can lead to increased resistance, or the virus can enter the cell and create an environment suitable for self-replication, resulting in secondary infection of neighboring and distant cells [30].
To identify transcriptomic changes common to glioma and NB cells upon infection with VV-GMCSF-Lact, we compiled lists of upregulated and downregulated transcripts in different cell cultures. By 12 h of infection, the vast majority of differentially expressed transcripts were culture-specific: 65.89% and 62.92% of unique upregulated and downregulated DEGs, respectively . Only the changes in the expression levels of five histone genes and the MARCKS gene could be considered common indicators of 12 h infection in all analyzed cell cultures and ). However, by 24 h of infection, only 38–39% of the DEGs remained unique to a particular cell culture. The levels of 85 mRNAs increased in all cell cultures, while the levels of 54 mRNAs were reduced and ). It should be noted that histone gene transcripts constituted a significant, almost definitive group among all common activated genes, at both 12 h and 24 h of infection , Figure 4, discussed below).
Considering that changes in individual RNA levels only partially reflect the complex and interconnected processes of the viral infection response, we analyzed common characteristics of the up/downregulated transcript sets using the Enrichr platform [21]. According to the Enrichr “ENCODE and ChEA Consensus TFs from ChIP-X” library, IRF (IRF1, IRF8), REST, FOSL2, SRF, and RELA (member of NF-kB family) are outlined as common regulators of activated human host genes in the process of VV-GMCSF-Lact infection, whereas NFY (NFYA and NFYB) and E2F (E2F1, E2F4 and E2F6) transcription factor control sets of human DEGs are suppressed following infection with the virus .
To describe common text annotations for up- or downregulated genes, a set of Enrichr libraries was selected, including GO, KEGG, MSigDB Hallmark, and Panther . Common annotations of upregulated transcript clusters include inflammatory and antiviral response pathways, such as cytokine-mediated signaling pathway (GO:0019221); defense response to virus (GO:0051607); chemokine and cytokine activities (GO:0008009, GO:0005125, respectively); NF-kappa B signaling pathway; inflammatory response; and others. Clusters of upregulated histone transcripts form separate groups in Enrichr terms: nucleosome organization (GO:0034728), DNA binding (GO:0003677), and viral carcinogenesis. In addition, annotations of downregulated transcript clusters include cell cycle-related nuclear processes: mitotic spindle organization (GO:0007052), microtubule cytoskeleton organization involved in mitosis (GO:1902850), G2-M checkpoint, cytoskeletal regulation via Rho GTPase, and others .
We compared sets of DEGs regulated by selected transcription factors and grouped by common text annotations . This enabled us to construct generalized schemes of activated (Figure 5) or suppressed (Figure 6) processes that are common for gliomas and NB cells upon infection with VV-GMCSF-Lact.
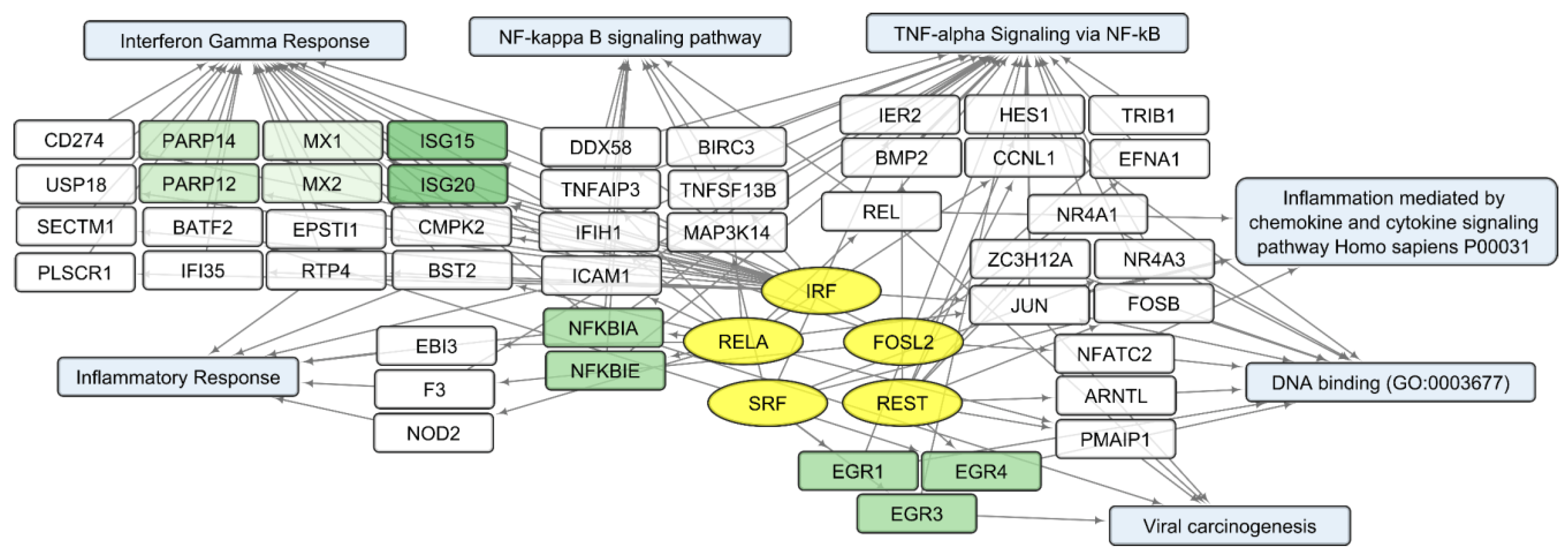
Figure 5. Scheme of the relationship between transcripts, transcription factors, and processes activated in glioma and NB cells upon infection with VV-GMCSF-Lact. Activated transcription factors IRF, RELA, FOSL2, SRF, and REST are shown in yellow ovals; selected activated genes—in white and green rectangles; and signaling pathways, biological processes, and gene annotations—in blue rectangles. Groups of genes with similar functions are drawn together using shades of green. Based on the analysis of the top 300 activated genes using Enrichr libraries: “ENCODE and ChEA Consensus TFs from ChIP-X”; “MSigDB Hallmark 2020”; “GO Biologic Process 2021”; “Panther 2016”; and “KEGG 2021 Human”.
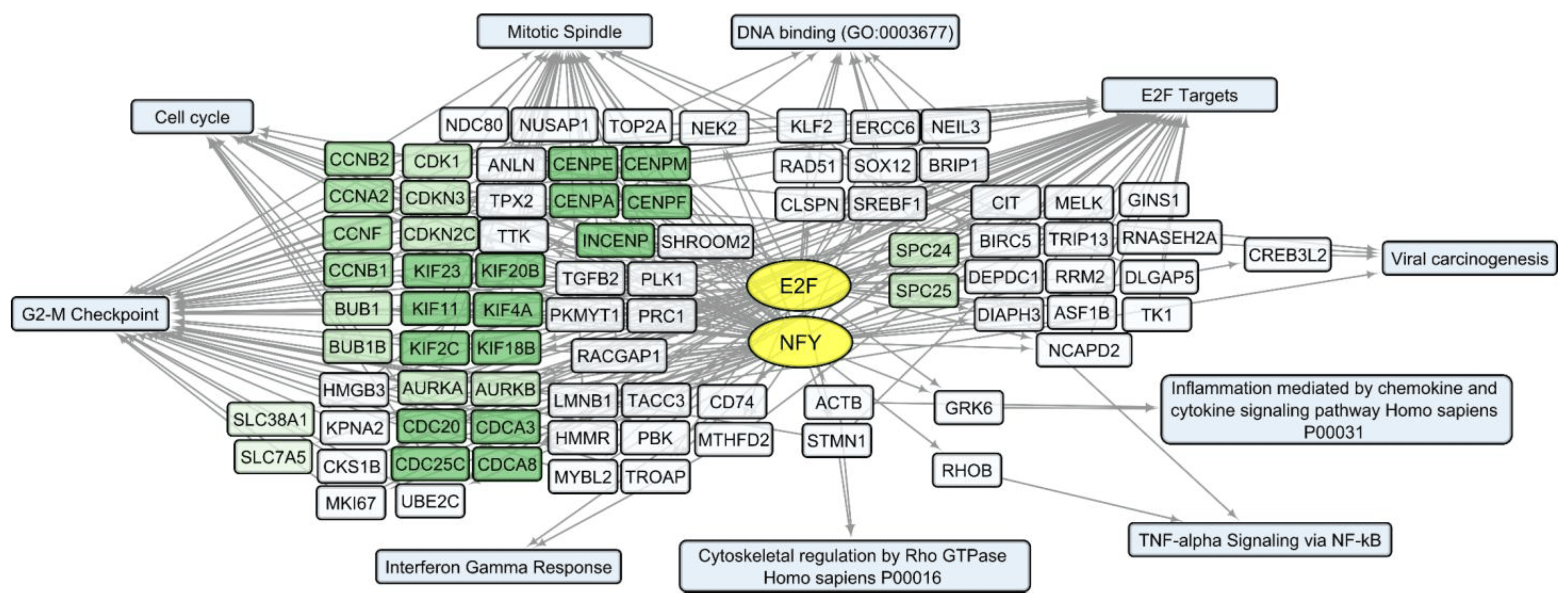
Figure 6. Scheme of the relationship between transcripts, transcription factors, and processes commonly suppressed in glioma and NB cells upon infection with VV-GMCSF-Lact. The suppressed transcription factors of the E2F family (E2F1, E2F4, and E2F6) and NFY family (NFYA and NFYB) are shown in yellow ovals; selected activated genes—in white and green rectangles; and signaling pathways, biological processes, and other gene annotations—in blue rectangles. Groups of genes with similar functions are drawn together using shades of green. Based on the analysis of the top 300 downregulated genes using Enrichr libraries: “ENCODE and ChEA Consensus TFs from ChIP-X”; “MSigDB Hallmark 2020”; “GO Biologic Process 2021”; “Panther 2016”; and “KEGG 2021 Human”.
Within the sets of upregulated DEGs common to glioma and NB cells, separate groups of genes can be further distinguished. Their products are involved in the antiviral, proinflammatory response (Figure 5). These are NFKBIA and NFKBIE, feedback inhibitors of the NF-kappa-B/REL transcription factor family [31]. Members of the Early Growth Response family of transcription regulators (EGR1, 3, and 4) play an important role in the regulation of cellular responses to growth factors, DNA damage, and viral infection. The induction of EGR1 following viral infection stimulates multiple inflammatory factors (EGR2 and EGR4) and mediates host cell response to viruses [32]. Products of ISG15 and ISG20 are functionally different proteins—a ubiquitin-like modifier and an exonuclease, respectively, although both ISG15 and ISG20 belong to interferon-stimulated genes and are involved in the innate immune response to viral infection [33].
Among the human genes, whose expression is commonly suppressed in glioma and NB cells upon infection with VV-GMCSF-Lact, several functionally related gene clusters can be distinguished (Figure 6). These are groups of genes encoding cyclins (CCNB1, CCNB2); cyclin-dependent kinases (CDK1, CDKN3, and CDKN2C); the Aurora subfamily of cell cycle-regulated protein kinases (AURKA and AURKB); proteins associated with the cycle of cell division (CDC20, CDCA8); motor proteins of the kinesin family required for mitosis (KIF11, KIF23); and genes of centromere proteins (CENPA, CENPF).
Taken together, our data indicate that infection with VV-GMCSF-Lact commonly induces the expression of genes encoding elements of the innate antiviral immune response in both glioma and NB cells while suppressing various genes involved in cell division and cell cycle regulation.
3.3. VV-GMCSF-Lact Transcripts in Infected Glioma and NB Cell Cultures
Early VACV gene expression begins shortly after infection and persists for 2–3 h. Early genes encode the proteins of virus uncoating, transcription regulation for the subsequent phase of intermediate gene expression, and various proteins needed for the replication of the viral genome [34]. The transcription of late genes, which encode late proteins that are necessary for the assembly of new virions, starts at ~6 h post-infection [35].
In this work, we performed an NGS-transcriptomic analysis of human brain cells 12 h and 24 h after infection with the VV-GMCSF-Lact virus (1 PFU/cell), so as to reveal “quasi-stationary” virus transcripts detected from the onset of infection to the death of the host cell. To describe major VV-GMCSF-Lact genes that are commonly expressed in glioma and NB cell cultures, we ranked viral transcripts by their relative contribution in each cell culture (12 h and 24 h time points combined). We compiled a list of the top 25 genes with generally high expression by averaging and sorting the ranks assigned to individual transcripts .
The list of VV-GMCSF-Lact transcripts expressed in both glioma cell cultures and NB cells includes products of genes that are characteristic of the VAVC “early”, “intermediate”, and “late” phases. Among the 25 commonly expressed viral genes, there are those encoding structural proteins, DNA polymerase components, host defense inhibitors, and polypeptides with other functions . Additionally, the distribution of these 25 genes in the VV-GMCSF-Lact genome does not show any significant shifts to the 5′- or 3′-ends of viral DNA (Figure 7). Thus, VV-GMCSF-Lact genes that are commonly expressed in infected human glioma and NB cells should be described within the specific context of the host cell transcriptome, rather than solely based on their location in the virus DNA or their annotated functions (see below for further discussion).
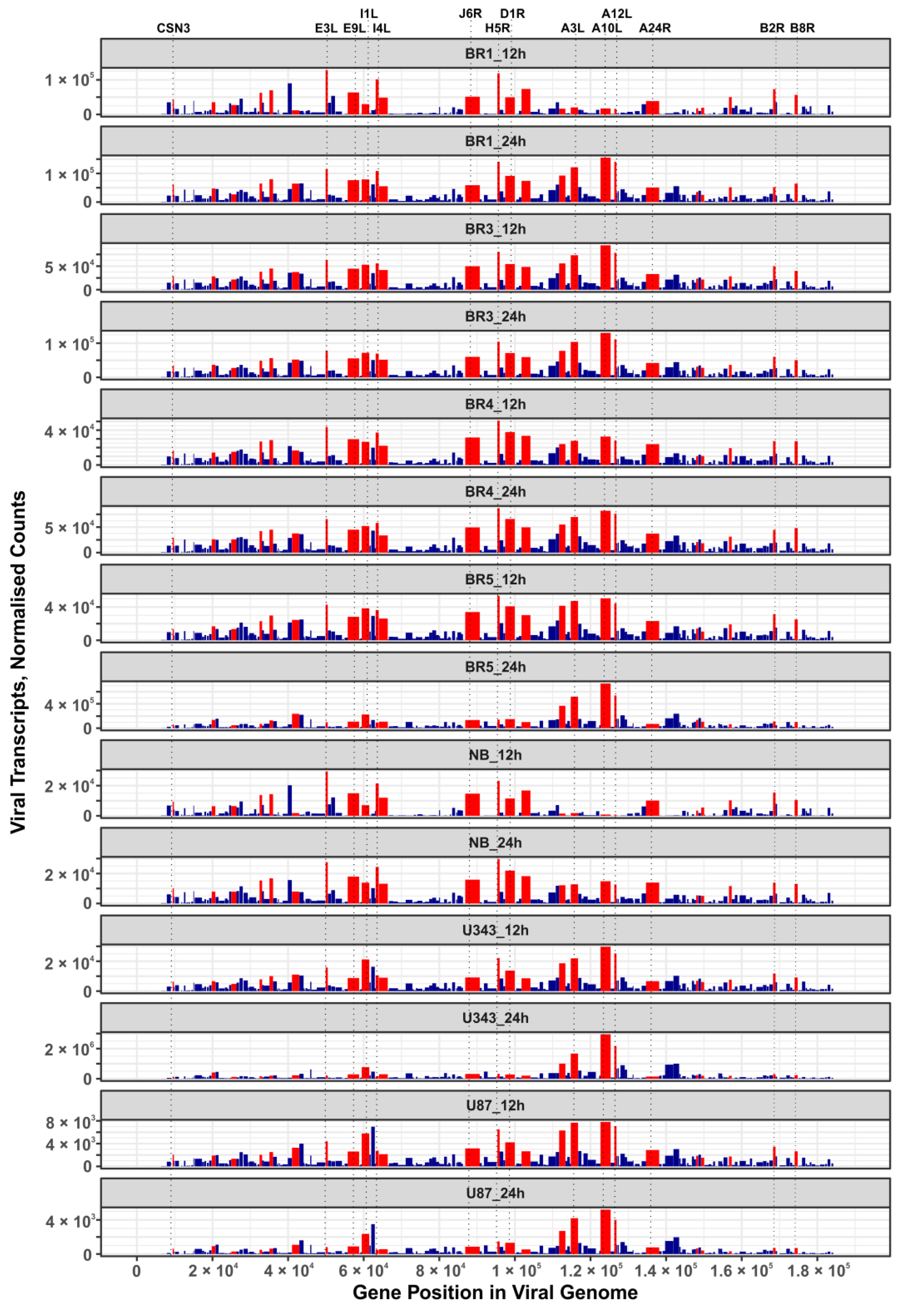
Figure 7. Expression maps of VV-GMCSF-Lact genes in glioma and NB cells. Bar plots display normalized DESeq2 RNA counts of VV-GMCSF-Lact transcripts (y-axis) across the length of the viral genome (x-axis). The width of the bars corresponds to the length of the gene in the virus genome. Red bars highlight the top 25 viral genes whose transcription is commonly increased in cells after 12 h and 24 h of infection . The vertical dotted lines indicate the positions of the selected viral genes in the VV-GMCSF-Lact genome.
It should also be noted that the CSN3 gene transcript encoded by VV-GMCSF-Lact (a fragment of the human CSN3 gene lactaptin, which we inserted into the parental VACV genome), was found among the 25 most commonly expressed viral transcripts and Figure 7).
3.4. Human Transcripts That Correlate with the VV-GMCSF-Lact RNA in Glioma and NB Cells
Apart from identifying common DEGs, another approach can help describe how cells respond to viral infection. It is based on the analysis of human transcripts whose levels correlate directly or inversely with the total level of viral transcripts. Having RNA sequencing data for the transcriptomes of seven cell cultures combined with the viral transcriptome data at each time point , we performed a correlation analysis to identify sets of human genome transcripts whose levels correlate with the total VV-GMCSF-Lact RNA in infected cells. Two sets of human genome transcripts were selected: 300 transcripts showing the best positive correlation with the number of virus transcripts and 300 transcripts with the best negative correlation (representative plots are shown in ).
The set of 300 human transcripts showing the strongest positive correlation with total VV-GMCSF-Lact RNA levels is enriched in transcripts regulated by such transcription factors as ATF2, BRCA1, CREB1, TAF1, and YY1 (Enrichr terms form “ENCODE and ChEA Consensus TFs from ChIP-X” library, ). The set of 300 negatively correlating transcripts is dominated by gene products regulated by the MYC-MAX family and the NFY family, as well as by the same transcription factors as in the case of positive correlation: ATF2, CREB1, TAF1, and YY1 .
Notably, some transcription factors (ATF2, CREB1, TAF1, and YY1) can control non-overlapping sets of genes that correlate positively or negatively with the total level of viral RNA . This is likely due to the well-known fact that viral transcription factors modulate gene expression by replacing or regulating the activity of host cell transcription machinery [34,59]. From this point of view, host cell transcription factors, which are influenced by the virus and induce both the activation and suppression of transcription in infected cells, can be considered the primary targets of viral transcriptional regulators. The diversity of transcription factors correlating directly or inversely with the level of viral RNA in human cells is summarized in Figure 8 and Figure 9, together with the corresponding processes and pathways.
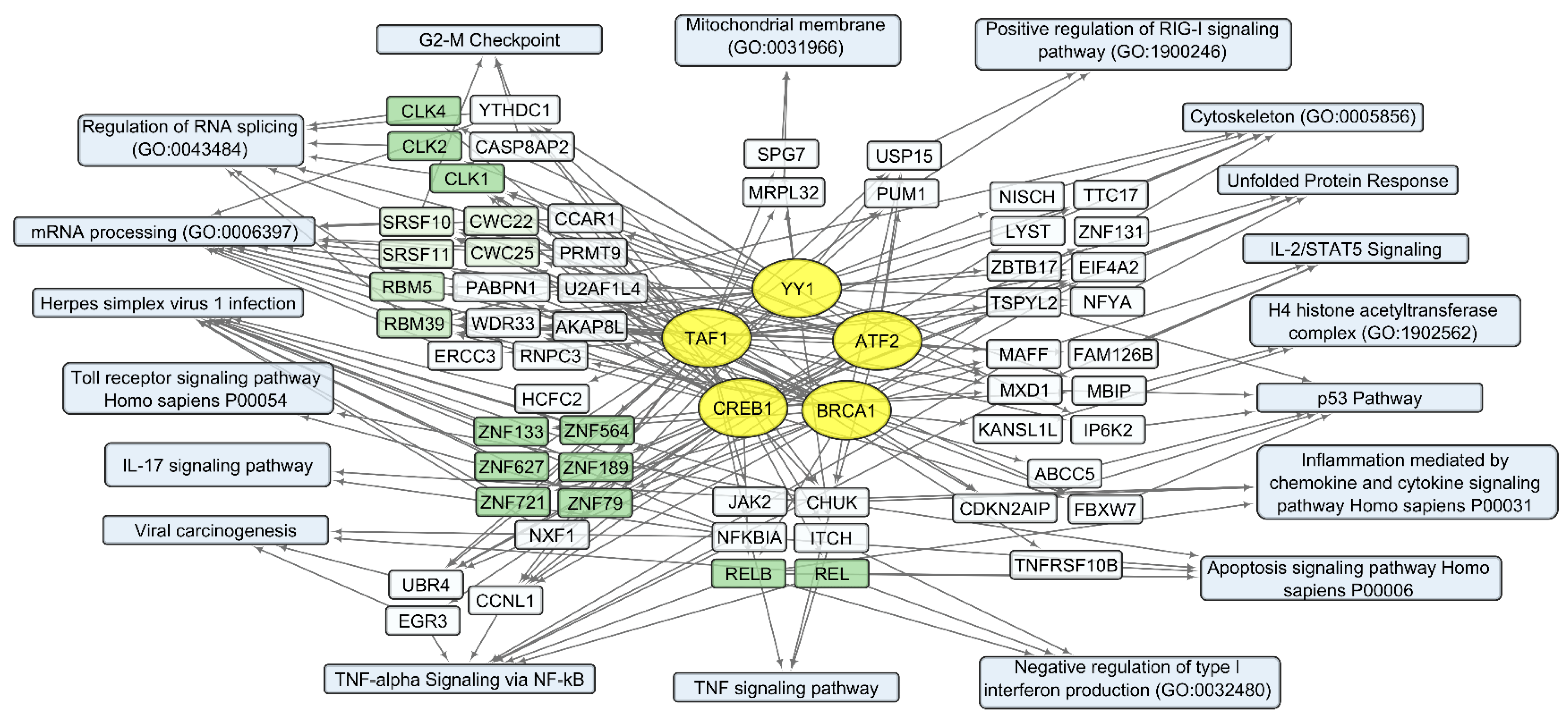
Figure 8. Human transcripts that positively correlate with the total VV-GMCSF-Lact RNA in glioma and NB cells. The scheme illustrates the relationships between the activity of transcription factors YY1, ATF2, BRCA1, CREB1, and TAF1 (yellow ovals) and transcripts that positively correlate with total VV-GMCSF-Lact RNA (white and green rectangles). The associated signaling pathways, biological processes, and other gene annotations are shown in blue rectangles. Groups of genes with similar functions are drawn together using shades of green based on the analysis of gene sets using the Enrichr libraries “ENCODE and ChEA Consensus TFs from ChIP-X”; “MSigDB Hallmark 2020”; “GO Biologic Process 2021”; “KEGG 2021 Human”; and “Panther 2016”.
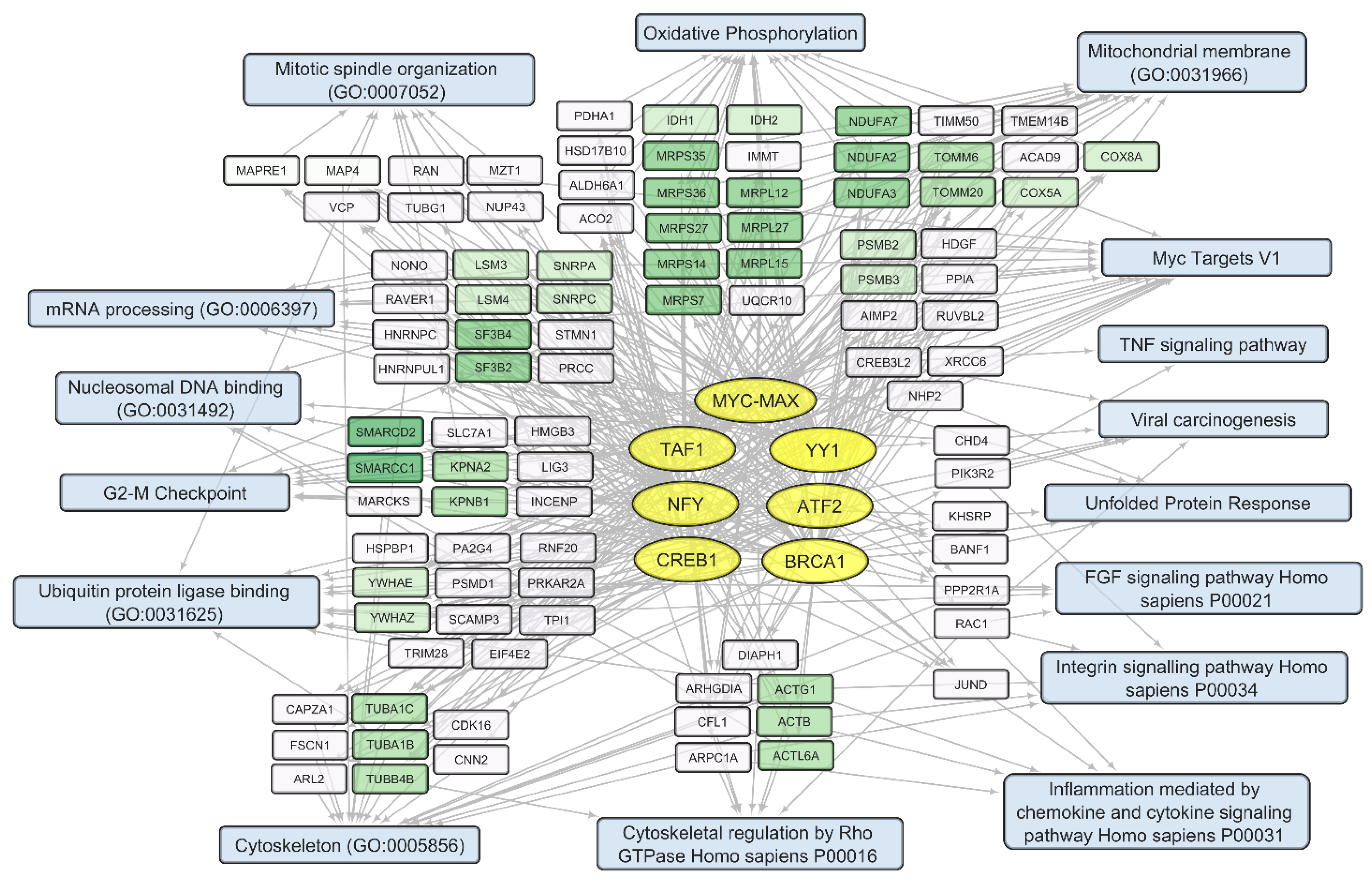
Figure 9. Human transcripts that negatively correlate with the total VV-GMCSF-Lact RNA in glioma and NB cells. The scheme illustrates the relationship between the activity of transcription factors of the MYC-MAX family, YY1, ATF2, BRCA1, CREB1, NFY family, and TAF1 (yellow ovals) and transcripts that negatively correlate with the total number of viral RNA reads (white and green rectangles). The associated signaling pathways, biological processes, and other gene annotations are shown in blue rectangles. Groups of genes with similar functions are drawn together using shades of green based on the analysis of gene sets using the Enrichr libraries “ENCODE and ChEA Consensus TFs from ChIP-X”; “MSigDB Hallmark 2020”; “GO Biologic Process 2021”; and “KEGG 2021 Human”.
Among human transcripts that correlate positively with viral RNA, histone gene products should be listed separately . Histone gene aliases are not uniformly represented in the Enrichr libraries and, therefore, are included in a separate row in . All the listed histone gene transcripts are present not only among mRNAs that positively correlate with the level of viral RNA but also within the sets of DEGs that are commonly found in infected glioma cells . Thus, histone gene transcripts are usually upregulated upon infection of glioma by VV-GMCSF-Lact and their relative level can reflect the “depth” of infection corresponding to the amount of viral RNA in cells.
One of the most significant groups of genes that positively correlate with viral RNA is “Herpes simplex virus 1 infection” (, Figure 8). This group of transcripts indicates the cellular response characteristic of infection with double-stranded DNA viruses, which include members of the Herpes virus family and VACV. Additionally, there are groups of genes encoding proteins that are involved in mRNA processing and regulation of RNA splicing: Cdc2-like protein kinases CLK1, −2, 4; RNA splicing factors SRSF10 and SRSF11; components of the spliceosome CWC22 and CWC25; and pre-mRNA splicing factors RBM5 and RBM39 (, Figure 8).
Among genes whose expression correlates inversely with the level of viral RNA, it is necessary to single out the groups encoding mitochondrial proteins. These are the components of mitochondrial ribosome proteins (MRPS and MRPL groups of genes) and mitochondrial membrane proteins (Isocitrate Dehydrogenase 2 IDH2; Ubiquinone Oxidoreductase Subunits NDUFA2, −3, −7; Cytochrome C Oxidase Subunit COX5A and COX8A; and others involved in oxidative phosphorylation). Genes encoding components of the cytoskeleton and its regulation are also worth noting. These include tubulins TUBA1C, TUBA1B, and TUBB4B and actins ACTB, ACTG1, and ACL6A. An increase in viral transcripts is accompanied by a downregulation of genes involved in the mitotic spindle organization and the G2/M checkpoint (and others in , Figure 9), which aligns with the groups described as commonly suppressed genes (, Figure 6).
Thus, an increase in the level of VV-GMCSF-Lact RNA in glioma and NB cells is accompanied by the transcriptional activation of histones, genes involved in post-transcriptional RNA processing, and genes associated with inflammation through chemokine and cytokine signaling pathways, apoptosis, etc. ( and Figure 8). Host genes whose expression is suppressed with an increase in the level of VV-GMCSF-Lact RNA encode mitochondrial proteins, cytoskeletal proteins, and cell cycle regulators at the mitotic spindle organization stage and the G2/M checkpoint (Figure 9, ).
3.5. Human Transcripts That Correlate with the VV-GMCSF-Lact Cytotoxic Dose CD50 in Glioma and NB Cells
One of the most intriguing questions regarding VV-GMCSF-Lact infection is the resistance/susceptibility of human cells to the cytotoxic effects of the virus. Therefore, we analyzed sets of human transcripts whose expression correlates directly or inversely with the cytotoxic dose of VV-GMCSF-Lact (CD50, . When the relative level of human mRNAs correlates directly with CD50, they could perhaps be considered as transcripts associated with the cell’s resistance to the cytotoxic effect of the virus. Similarly, mRNAs that correlate inversely with CD50 might be linked to processes mediating the cell’s sensitivity to the cytotoxic effect of VV-GMCSF-Lact .
Transcripts of VV-GMCSF-Lact-infected glioma and NB cells that directly correlate with CD50 are enriched with mRNAs controlled by transcription factors ATF2, BRCA1, CREB1, ELF1, TAF1, UBTF, and YY1. The sets of transcripts that inversely correlate with CD50 are enriched with gene products controlled by the MYC-MAX, E2F, and NFY families of transcription factors, as well as by BRCA1, CREB1, TAF1, and YY1 . BRCA1, CREB1, TAF1, and YY1 are the transcription factors that control both sets of genes, i.e., those that either increase or decrease their expression in correlation with the level of the virus RNA . Moreover, as in the case of the transcripts correlating with the viral RNA level , sets of mRNAs that correlate positively and negatively with CD50 are enriched with those controlled by a similar set of transcription factors (BRCA1, CREB1, TAF1, and YY1; ). Considering that during infection, the transcription of human genes is significantly modulated by viral factors [34,59], it can be expected that the gene enrichment analysis shows multidirectional changes in the activity of most affected host cell transcription factors. Importantly, the Enrichr library “ENCODE and ChEA Consensus TFs from ChIP-X” does not include patterns of the genes targeted by viral transcription factors/modulators. Therefore, the action of the viral transcription factors manifests through a combination of the Enrichr library tabulated human TFs.
Downregulated mRNAs common to glioma and NB cells are enriched in transcripts controlled by transcription factors of the E2F and NFY families . A decrease in the activity of MYC-MAX and E2F suggests the suppression of human genes with an increasing level of viral RNA . The MYC-MAX, E2F, and NFY families of transcription factors are associated with the sensitivity of cells to the cytotoxic effect of the virus . Thus, it can be assumed that VV-GMCSF-Lact infection proceeds with the suppression of E2F and NFY, while the decrease in MYC-MAX and E2F activity is directly related to the increase in viral transcripts. The decreased activity of all three families, MYC-MAX, E2F, and NFY, is linked to the sensitivity of cells to the cytotoxic effect of the virus .
Among biological processes and pathways involving genes whose expression positively correlates with the VV-GMCSF-Lact CD50 , the following are distinguished: a group of small RAB GTPases (RAB1A, RAB2A), regulators of intracellular membrane trafficking; components of the coat protein complex II, which promotes transport vesicle generation from the endoplasmic reticulum (SEC24B, SEC31A); vesicular coat proteins (COPA, COPB1, and COPB2); and dynactins (DCTN1, 4–6) that regulate the transport of vesicles and organelles along microtubules (summarized in Figure 10).
Figure 1. Representative images of immortalized human glioblastoma lines (U343, U87), NB cells, and patient-derived glioma cell cultures (BR1, BR3, BR4, BR5).
Negative correlations with the VV-GMCSF-Lact CD50 can be observed within several functionally related groups (, Figure 11). These encode mitochondrial ribosomal proteins (MRPL9, MRPS15); mitochondrial cytochrome c oxidase subunits COX10, COX7C; and mitochondrial respiratory chain complex I—NADH:ubiquinone oxidoreductase subunits (NDUFAB1, NDUFB3). Additionally, there are genes involved in the processing of pol II transcripts: SRSF encoding the processing and regulation of RNA splicing (RNA splicing factors SRSF1, −2, −3); heterogeneous nuclear ribonucleoproteins HNRNPD, HNRNPA0; small nuclear ribonucleoproteins SNRPA, SNRPA1; transcription and splicing regulators RBM10, −14, −15; and others (, Figure 11).
Taken together, the data on the correlation of human transcript levels with the VV-GMCSF-Lact CD50 emphasize that the resistance of glioma and NB cells to viral infection is largely associated with the activity of genes encoding components of endoplasmic reticulum, intracellular transport, and secretion. The susceptibility of cells to viral cytotoxicity is associated with the activity of human genes encoding mitochondrial proteins and factors of nuclear RNA processing and splicing.
3.6. Viral Transcripts That Correlate with the Cytotoxic Dose of the Virus in Glioma and NB Cells Infected with VV-GMCSF-Lact
The search for mRNAs expressed in correlation with the cytotoxic dose applies to both human and viral genome transcripts. Therefore, we analyzed the correlation between the viral CD50 values and the levels of viral transcripts in VV-GMCSF-Lact-infected glioma and NB cells. No viral mRNAs correlate positively with CD50 (Pearson R > 0.5). This suggests that none of the VV-GMCSF-Lact transcripts are associated with processes providing cellular resistance to the cytotoxic effects of the virus.
Among the VV-GMCSF-Lact transcripts that inversely correlate with CD50 and are thus associated with its cytotoxicity, the following stand out: H5R (late transcription factor), A24R (VLTF-4 DNA-dependent RNA polymerase subunit rpo132), and E3L (double-strand RNA-binding protein). H5R, A24R, and E3L are also detected as commonly expressed transcripts . Importantly, the level of viral CSN3 RNA negatively correlates with the VV-GMCSF-Lact CD50 in glioma and NB cells, ranking second (R = −0.654) after the viral H5R transcript R = −0.714, Figure 12).
Thus, the cytotoxicity of VV-GMCSF-Lact against glioma and NB cells results from both VACV genome-encoded factors and modifications to the viral genome by the human CSN3 gene fragment encoding the proapoptotic peptide lactaptin.
4. Discussion
Oncolytic virotherapy is a rapidly evolving approach that aims to selectively kill cancer cells while leaving normal, non-cancerous cells viable. Vaccinia virus (VACV) is one of the most explored platforms for creating oncolytic treatments for various malignant neoplasms [60]. Vaccinia is a dsDNA virus of the Poxviridae family. Its DNA (~195 kb) encodes approximately 250 genes [34] and, together with proteins and lipids, forms a ~360 nm × 270 nm × 250 nm virion [44]. The virus enters cells via either micropinocytosis or direct fusion with the plasma membrane [61]. The infection is accompanied by a global change in the molecular and genetic landscape of the cell. It can lead to either the suppression of the virus or the death of the infected cell. A previous study using deep RNA sequencing on VACV-infected cervical cancer Hela cells identified upregulated host cell RNAs linked to the NF-κB cascade, apoptosis, signal transduction, and ligand-mediated signaling, likely representing the response to the virus invasion [62].
Recently, we developed a recombinant VACV named VV-GMCSF-Lact as a carrier for creating drugs for the treatment of malignant tumors, including glioblastoma. To construct VV-GMCSF-Lact from the original L-IVP strain, the viral thymidine kinase (tk) gene was replaced with the human GM-CSF, and the viral growth factor (vgf) gene was replaced with a fragment of the human CSN3 gene encoding the proapoptotic peptide lactaptin [11]. Here, we applied NGS to analyze the effect of VV-GMCSF-Lact infection on the transcriptome of human glioma and normal brain cells in culture . In terms of the sample tree and PCA, VV-GMCSF-Lact infection induced significant transcript changes in all analyzed cell cultures (Figure 3). The number of differentially expressed human genes correlated with the relative amount of viral transcripts . Thus, the data allowed us to describe trends in viral infection, covering both general and cell culture-specific patterns. To overview the effect of VV-GMCSF-Lact on cells, we analyzed sets of transcripts that are typically up/downregulated in infected cells, exhibit positive or negative correlations with the viral RNA level in cells, or are associated with the virus cytotoxic dose (CD50).
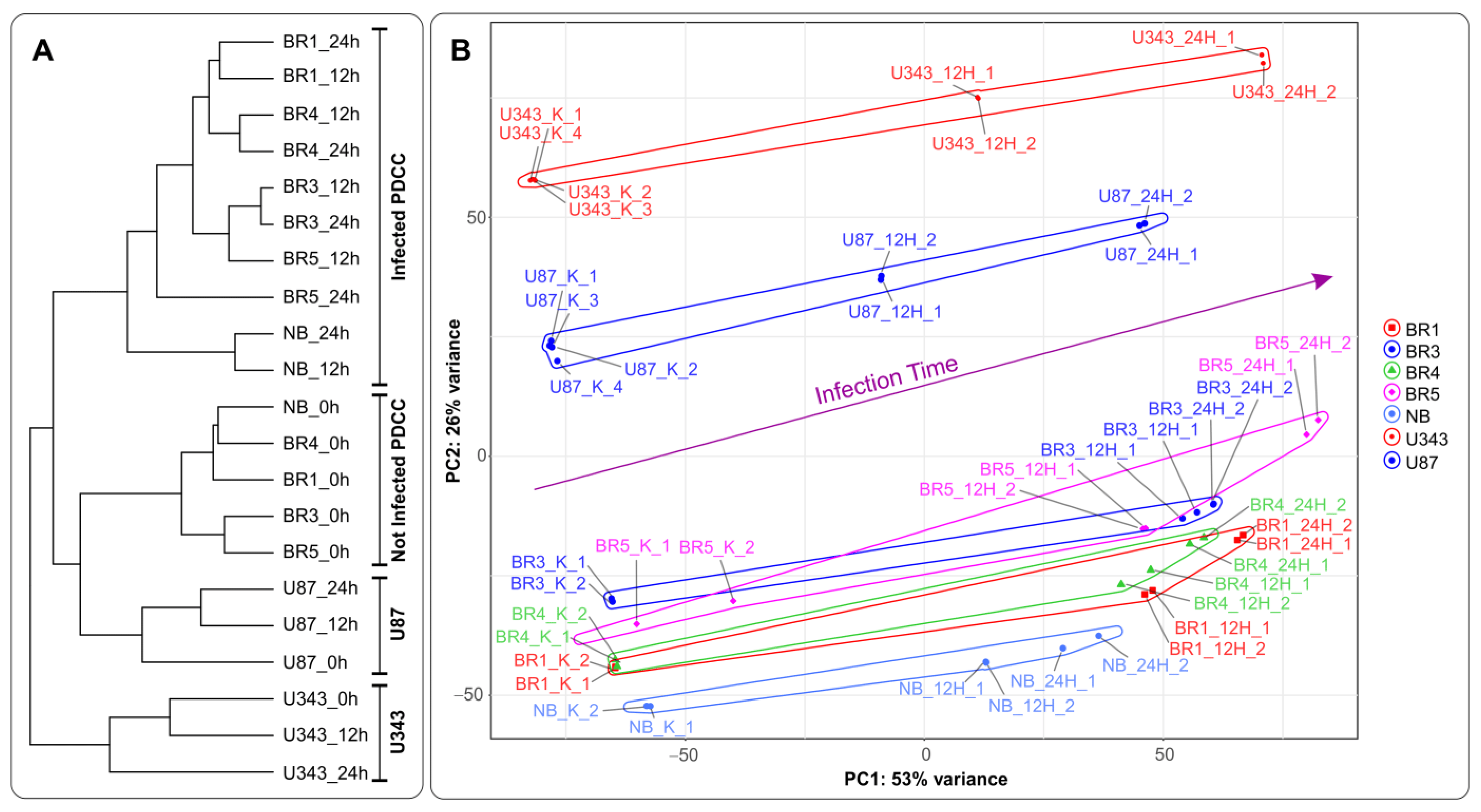
Figure 3. (A) A tree of Euclidean distances of variance stabilizing transformed (VST) RNA expression data of immortalized glioblastoma, patient-derived glioma cell cultures, and NB cells. The complete agglomeration method was used for clustering. (B) Principal component analysis of DESeq2 normalized VST-transformed RNA expression data. Sample-specific PC1:PC2 points are annotated with cell-defined envelopes. The deep purple arrow shows the common trend of PC1:PC2 transition from noninfected (0 h) to the 12 h and 24 h VV-GMCSF-Lact-infected state. Patient-derived cell cultures (PDCCs).
Among human transcripts that are commonly upregulated following VV-GMCSF-Lact infection, special attention should be given to mRNA of histone genes. Transcripts representative of all major histone families (H1/H5, H2A, H2B, H3, and H4) are either generally activated upon infection or correlate positively with the total viral RNA levels in glioma and NB cells (Figure 4, and ). This aligns with data from the literature, as activation of histone gene expression is also observed upon infection of Macaca mulatta kidney epithelial cells with another member of the Poxviridae family, the Monkeypox virus [63,64].
It is known that double-stranded VACV DNA is replicated in cytoplasmic structures [65], and cellular proteins such as histones are not involved in VACV genome organization [34]. If present in viral factories, histones are not directly involved in the life cycle of the virus. Therefore, we should not assume that histones interact with viral DNA, even though the transport/exchange of proteins between the cytoplasmic viral factories and the nucleus has been shown for viral proteins [46,66]. However, it can be hypothesized that the activation of histone genes induced by VV-GMCSF-Lact reflects the modulation of nuclear processes, including nucleosome organization, mitotic spindle organization, and the G2/M checkpoint in particular (, Figure 4). The upregulation of histone genes may result from the viral factors affecting nuclear DNA compaction and, in general, from the suppression of the cell cycle of an infected cell. This is indirectly confirmed by data indicating the involvement of the VACV K7 protein in histone methylation [67] and the C6 protein in proteasomal degradation of histone deacetylase [68].
Transcripts commonly downregulated during VV-GMCSF-Lact infection and showing negative correlation with total viral RNA in glioma and NB cells are enriched in mRNA of genes that regulate processes like the mitotic spindle organization, mitotic sister chromatid segregation, the G2/M checkpoint, and other cell cycle-related functions. The activity of genes controlling vital processes such as cytoskeletal function, mitochondrial translation, and oxidative phosphorylation is also suppressed upon the infection (, Figure 6 and Figure 9). Thus, VV-GMCSF-Lact infection significantly modulates nuclear processes, redirecting them to the inhibition of cellular DNA replication, cell cycle arrest, and the suppression of vital cytoplasmic and mitochondrial pathways. Moreover, genes of the antiviral inflammatory response are activated in infected glioma and NB cells. These include groups of transcripts that mediate a defense response to the virus: interferon-gamma response, NF-kappa B signaling pathway, inflammation mediated by chemokine and cytokine signaling pathways, and others (Figure 5, ). This is partly consistent with previous studies of the transcriptome of VACV-infected HeLa cells [62]. Overall, our results and data from the literature point to the competition between virus-induced suppression of cell viability and the host antiviral response to infection.
To understand what processes are responsible for the cellular resistance or vulnerability to the cytotoxic effect of VV-GMCSF-Lact, we analyzed the correlation between the levels of individual cellular RNAs of glioma and NB cells with the CD50 cytotoxic dose of the virus. Transcript groups that show positive correlations with CD50 and that are thus are associated with cellular resistance to viral cytotoxicity encode gene products involved in protein transport, protein secretion, and the endoplasmic reticulum to Golgi vesicle-mediated transport (, Figure 11).
It is known that the life cycle of VACV begins in the plasma membrane. The replication of the vaccinia virus occurs in the cytoplasmic crescent-shaped vesicle-like structures, resembling mini-nuclei, which originate from the smooth endoplasmic reticulum (“viral factories” [69]). Both viral and cellular proteins move between viral factories and the host cytoplasm [65]. The synthesis and assembly of VACV particles relies on the host’s translational apparatus and protein transport occurring between the endoplasmic reticulum and the Golgi apparatus (reviewed in [59]). Our data suggest that glioma and NB cells are less vulnerable to the cytotoxic effect of VV-GMCSF-Lact due to increased expression of the genes of the RAB, DCTN, SEC, and COPA families, which encode the components of Golgi vesicles, protein transport, and secretion (Figure 10, ).
Important gene clusters, whose expression correlates negatively with the VV-GMCSF-Lact CD50, encode proteins from the SR, RBM, SNRP, HNRNP, and NUP families, which participate in the maturation of pol II nuclear transcripts and mRNA splicing (, Figure 11). The VACV genes lack introns, and their transcription is not thought to require the nuclear apparatus for mRNA splicing [70]. The decapping of transcripts from intron-containing genes helps the virus to deplete host transcripts and remodel the infected cell transcriptome [71]. Huang et al. showed that in VACV-infected human cells, SR proteins are hypophosphorylated and functionally inactivated as splicing repressors or splicing enhancers [72]. These data indicate that VACV infection represses post-transcriptional processing and modulates the nuclear-cytoplasmic transport of pol II transcripts in the host cell. Considering our data on the negative correlation of human cell transcripts with the VV-GMCSF-Lact CD50 (, Figure 11), it can be proposed that an upregulation of genes that control mRNA splicing and processing of capped intron-containing pre-mRNAs represents one of the main factors of cell sensitivity to the cytotoxic effect of the virus.
Genes showing a positive or negative correlation with CD50 may be of interest for developing targeted drugs aimed at enhancing the effectiveness of virotherapy (, Figure 10 and Figure 11). For example, by knocking down genes whose expression correlates positively with CD50, one can expect an increase in the sensitivity of tumor cells to the virus. A similar effect could be promoted by stimulating the expression of genes that inversely correlate with CD50.
Among the viral transcripts that are commonly expressed in VV-GMCSF-Lact-infected glioma and NB cells, there is E3L, encoding a dsRNA-binding protein involved in the inhibition of innate immune responses; E9L, the catalytic subunit of viral DNA polymerase; I3L, an ssDNA-binding protein; I4L, the ribonucleotide reductase large subunit; and CSN3, or lactaptin, the fragment of human kappa-casein gene inserted in the VV-GMCSF-Lact genome . This suggests that our VV-GMCSF-Lact viral construct is a powerful vector for the expression of genes in human cells. It is noteworthy that viral CSN3 mRNA is the second most important transcript (after VACV H5R) showing a negative correlation with CD50, as it is directly related to the cytotoxic effect of the virus on glioma cells (Figure 12). These data indicate that the cytotoxicity of the VV-GMCSF-Lact construct against glioma cells is mediated by both vaccinia virus genes and the recombinant gene encoding the human proapoptotic peptide lactaptin.
Overall, our study links the activity of transcription factors and their target genes to the cellular processes and signaling pathways involved in VV-GMCSF-Lact infection of glioma and NB cells. The list of transcription factors modulated by the infection includes the IRF, MYC-MAX, NFY, and E2F families; RELA; YY1; CREB1; and others (; Figure 5, Figure 6, Figure 8, Figure 9, Figure 10 and Figure 11). To describe these transcription factors, we compiled a brief review of data from the literature on their known properties, targets, and interactions .
Taken together, our data, highlighting transcription factors, molecular markers, biological pathways, and networks involved in glioma cell sensitivity to VV-GMCSF-Lact, can be used to improve the effectiveness of cancer virotherapy.
5. Conclusions
As one of the most extensively studied viral construct platforms, VACV has been explored for developing oncolytic therapy against various malignant neoplasms, including glioblastoma. Here, we describe the impact of a VACV-based viral construct, VV-GMCSF-Lact, on the transcriptomes of patient-derived glioma/glioblastoma cell cultures, immortalized human glioblastoma cells, and a culture of normal, non-malignant brain cells. For the first time, we conducted a thorough correlation analysis of the human transcriptomic data with viral RNA and the cytotoxic dose (CD50).
Both common and specific transcriptome changes indicate that mRNAs representing all major histone families are generally upregulated during infection with VV-GMCSF-Lact. They show positive correlations with viral RNA levels in glioma and NB cells. Genes of the antiviral inflammatory response are also activated in glioma and NB cells.
Conversely, genes involved in mitosis, the G2/M checkpoint, and other cell cycle-related functions are downregulated upon infection, and their transcripts correlate negatively with total viral RNA. The activity of genes controlling vital cellular processes, such as cytoskeletal function, mitochondrial translation, and oxidative phosphorylation, is also downregulated during the VV-GMCSF-Lact infection.
VV-GMCSF-Lact expresses the human CSN3 mRNA fragment encoding the proapoptotic peptide lactaptin. It is directly associated with the cytotoxic effect of the virus on glioma cells, once again affirming the rationale behind selecting this specific gene for insertion into the vector.
To identify RNA groups that are associated with cellular resistance or susceptibility to the cytotoxic effects of VV-GMCSF-Lact, we analyzed a correlation between individual transcripts in infected cells and the cytotoxic dose of the virus. Greater resistance to the cytotoxic effect of VV-GMCSF-Lact is observed in cells with the increased expression of genes encoding components of Golgi vesicles, protein transport, and secretion. In contrast, cells that are more sensitive to virus cytotoxicity demonstrate the upregulation of genes encoding proteins involved in the maturation of pol II nuclear transcripts and mRNA splicing. Notably, genes whose expression correlates positively or negatively with CD50 may be of interest for developing targeted drugs that increase the effectiveness of virotherapy. In conclusion, our data generalize the responses of glioma cells to the oncolytic virus infection and provide insight into the associated molecular pathways. These findings can be used to improve the effectiveness of virotherapy against malignant neoplasms.
References
- Ostrom, Q.T.; Patil, N.; Cioffi, G.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2013–2017. Neuro. Oncol. 2020, 22, iv1–iv96. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Li, X.; Sander, M.; Zhang, H.; Yan, G.; Lin, Y. Oncolytic Viro-Immunotherapy: An Emerging Option in the Treatment of Gliomas. Front. Immunol. 2021, 12, 721830. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K.; Núñez, F.J.; Haase, S.; McClellan, B.L.; Faisal, S.M.; Carney, S.V.; Yu, J.; Alghamri, M.S.; Asad, A.S.; Candia, A.J.N.; et al. Current Approaches for Glioma Gene Therapy and Virotherapy. Front. Mol. Neurosci. 2021, 14. [Google Scholar] [CrossRef]
- Todo, T.; Ito, H.; Ino, Y.; Ohtsu, H.; Ota, Y.; Shibahara, J.; Tanaka, M. Intratumoral Oncolytic Herpes Virus G47∆ for Residual or Recurrent Glioblastoma: A Phase 2 Trial. Nat. Med. 2022, 28, 1630–1639. [Google Scholar] [CrossRef] [PubMed]
- Thorne, S.; Bartlett, D.; Kirn, D. The Use of Oncolytic Vaccinia Viruses in the Treatment of Cancer: A New Role for an Old Ally? Curr. Gene Ther. 2005, 5, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Beerli, C.; Yakimovich, A.; Kilcher, S.; Reynoso, G.V.; Fläschner, G.; Müller, D.J.; Hickman, H.D.; Mercer, J. Vaccinia Virus Hijacks EGFR Signalling to Enhance Virus Spread through Rapid and Directed Infected Cell Motility. Nat. Microbiol. 2019, 4, 216–225. [Google Scholar] [CrossRef]
- Parato, K.A.; Breitbach, C.J.; Le Boeuf, F.; Wang, J.; Storbeck, C.; Ilkow, C.; Diallo, J.S.; Falls, T.; Burns, J.; Garcia, V.; et al. The Oncolytic Poxvirus JX-594 Selectively Replicates in and Destroys Cancer Cells Driven by Genetic Pathways Commonly Activated in Cancers. Mol. Ther. 2012, 20, 749–758. [Google Scholar] [CrossRef]
- Hengstschläger, M.; Knöfler, M.; Müllner, E.W.; Ogris, E.; Wintersberger, E.; Wawra, E. Different Regulation of Thymidine Kinase during the Cell Cycle of Normal versus DNA Tumor Virus-Transformed Cells. J. Biol. Chem. 1994, 269, 13836–13842. [Google Scholar] [CrossRef]
- Thorne, S.H.; Hwang, T.-H.H.; O’Gorman, W.E.; Bartlett, D.L.; Sei, S.; Kanji, F.; Brown, C.; Werier, J.; Cho, J.-H.; Lee, D.-E.; et al. Rational Strain Selection and Engineering Creates a Broad-Spectrum, Systemically Effective Oncolytic Poxvirus, JX-963. J. Clin. Investig. 2007, 117, 3350–3358. [Google Scholar] [CrossRef]
- Kochneva, G.; Sivolobova, G.; Tkacheva, A.; Grazhdantseva, A.; Troitskaya, O.; Nushtaeva, A.; Tkachenko, A.; Kuligina, E.; Richter, V.; Koval, O. Engineering of Double Recombinant Vaccinia Virus with Enhanced Oncolytic Potential for Solid Tumor Virotherapy. Oncotarget 2016, 7, 74171–74188. [Google Scholar] [CrossRef]
- Koval, O.A.; Tkachenko, A.V.; Fomin, A.S.; Semenov, D.V.; Nushtaeva, A.A.; Kuligina, E.V.; Zavjalov, E.L.; Richter, V.A. Lactaptin Induces P53-Independent Cell Death Associated with Features of Apoptosis and Autophagy and Delays Growth of Breast Cancer Cells in Mouse Xenografts. PLoS ONE 2014, 9, e93921. [Google Scholar] [CrossRef]
- Vasileva, N.; Ageenko, A.; Dmitrieva, M.; Nushtaeva, A.; Mishinov, S.; Kochneva, G.; Richter, V.; Kuligina, E. Double Recombinant Vaccinia Virus: A Candidate Drug against Human Glioblastoma. Life 2021, 11, 1084. [Google Scholar] [CrossRef]
- Hughes, L.; Wilkins, K.; Goldsmith, C.S.; Smith, S.; Hudson, P.; Patel, N.; Karem, K.; Damon, I.; Li, Y.; Olson, V.A.; et al. A Rapid Orthopoxvirus Purification Protocol Suitable for High-Containment Laboratories. J. Virol. Methods 2017, 243, 68–73. [Google Scholar] [CrossRef]
- Chou, T.-C. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Jurka, J.; Kapitonov, V.V.; Pavlicek, A.; Klonowski, P.; Kohany, O.; Walichiewicz, J. Repbase Update, a Database of Eukaryotic Repetitive Elements. Cytogenet. Genome Res. 2005, 110, 462–467. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Hartley, S.W.; Mullikin, J.C. QoRTs: A Comprehensive Toolset for Quality Control and Data Processing of RNA-Seq Experiments. BMC Bioinform. 2015, 16. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Caragher, S.; Chalmers, A.J.; Gomez-Roman, N. Glioblastoma’s Next Top Model: Novel Culture Systems for Brain Cancer Radiotherapy Research. Cancers 2019, 11, 44. [Google Scholar] [CrossRef]
- Da Hora, C.C.; Schweiger, M.W.; Wurdinger, T.; Tannous, B.A. Patient-Derived Glioma Models: From Patients to Dish to Animals. Cells 2019, 8. [Google Scholar] [CrossRef]
- Mooney, K.L.; Choy, W.; Sidhu, S.; Pelargos, P.; Bui, T.T.; Voth, B.; Barnette, N.; Yang, I. The Role of CD44 in Glioblastoma Multiforme. J. Clin. Neurosci. 2016, 34, 1–5. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, K.; Wang, Z.; Zhao, M.; Deng, Y.; Ji, W.; Zou, Y.; Qian, C.; Liu, Y.; Xiao, H.; et al. CD44-Mediated Poor Prognosis in Glioma Is Associated With M2-Polarization of Tumor-Associated Macrophages and Immunosuppression. Front. Surg. 2021, 8, 775194. [Google Scholar] [CrossRef]
- Zeng, J.; Xi, S.-Y.; Wang, F.; Liao, H.-D.; Yang, Y.-Z.; Hu, W.-M. L1CAM High Expression Associates with Poor Prognosis in Glioma but Does Not Correlate with C11orf95-RELA Fusion. Biomed Res. Int. 2020, 2020, 1353284. [Google Scholar] [CrossRef]
- Wachowiak, R.; Krause, M.; Mayer, S.; Peukert, N.; Suttkus, A.; Müller, W.C.; Lacher, M.; Meixensberger, J.; Nestler, U. Increased L1CAM (CD171) Levels Are Associated with Glioblastoma and Metastatic Brain Tumors. Medicine 2018, 97, e12396. [Google Scholar] [CrossRef]
- Maness, P.F.; Schachner, M. Neural Recognition Molecules of the Immunoglobulin Superfamily: Signaling Transducers of Axon Guidance and Neuronal Migration. Nat. Neurosci. 2007, 10, 19–26. [Google Scholar] [CrossRef]
- Dzwonek, J.; Wilczynski, G.M. CD44: Molecular Interactions, Signaling and Functions in the Nervous System. Front. Cell. Neurosci. 2015, 9, 175. [Google Scholar] [CrossRef]
- Burrell, C.J.; Howard, C.R.; Murphy, F.A. Pathogenesis of Viral Infections and Diseases. In Fenner’s Veterinary Virology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 47–78. ISBN 9780123751584. [Google Scholar]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Lehman, C.W.; Smith, A.; Kelly, J.; Jacobs, J.L.; Dinman, J.D.; Kehn-Hall, K. EGR1 Upregulation during Encephalitic Viral Infections Contributes to Inflammation and Cell Death. Viruses 2022, 14, 1210. [Google Scholar] [CrossRef]
- Schoggins, J.W. Interferon-Stimulated Genes: What Do They All Do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef]
- Greseth, M.D.; Traktman, P. The Life Cycle of the Vaccinia Virus Genome. Annu. Rev. Virol. 2022, 9, 239–259. [Google Scholar] [CrossRef]
- Sodeik, B.; Doms, R.W.; Ericsson, M.; Hiller, G.; Machamer, C.E.; Van’t Hof, W.; Van Meer, G.; Moss, B.; Griffiths, G. Assembly of Vaccinia Virus: Role of the Intermediate Compartment between the Endoplasmic Reticulum and the Golgi Stacks. J. Cell Biol. 1993, 121, 521–542. [Google Scholar] [CrossRef] [PubMed]
- Dueck, K.J.; Hu, Y.; Chen, P.; Deschambault, Y.; Lee, J.; Varga, J.; Cao, J. Mutational Analysis of Vaccinia Virus E3 Protein: The Biological Functions Do Not Correlate with Its Biochemical Capacity to Bind Double-Stranded RNA. J. Virol. 2015, 89, 5382–5394. [Google Scholar] [CrossRef] [PubMed]
- Bersch, B.; Tarbouriech, N.; Burmeister, W.P.; Iseni, F. Solution Structure of the C-Terminal Domain of A20, the Missing Brick for the Characterization of the Interface between Vaccinia Virus DNA Polymerase and Its Processivity Factor. J. Mol. Biol. 2021, 433, 167009. [Google Scholar] [CrossRef] [PubMed]
- Harrison, M.L.; Desaulniers, M.A.; Noyce, R.S.; Evans, D.H. The Acidic C-Terminus of Vaccinia Virus I3 Single-Strand Binding Protein Promotes Proper Assembly of DNA-Protein Complexes. Virology 2016, 489, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Gammon, D.B.; Gowrishankar, B.; Duraffour, S.; Andrei, G.; Upton, C.; Evans, D.H. Vaccinia Virus-Encoded Ribonucleotide Reductase Subunits Are Differentially Required for Replication and Pathogenesis. PLoS Pathog. 2010, 6, e1000984. [Google Scholar] [CrossRef]
- Mossman, K.; Upton, C.; Buller, R.M.; McFadden, G. Species Specificity of Ectromelia Virus and Vaccinia Virus Interferon-Gamma Binding Proteins. Virology 1995, 208, 762–769. [Google Scholar] [CrossRef]
- D’Costa, S.M.; Bainbridge, T.W.; Kato, S.E.; Prins, C.; Kelley, K.; Condit, R.C. Vaccinia H5 Is a Multifunctional Protein Involved in Viral DNA Replication, Postreplicative Gene Transcription, and Virion Morphogenesis. Virology 2010, 401, 49–60. [Google Scholar] [CrossRef]
- Schweneker, M.; Lukassen, S.; Späth, M.; Wolferstätter, M.; Babel, E.; Brinkmann, K.; Wielert, U.; Chaplin, P.; Suter, M.; Hausmann, J. The Vaccinia Virus O1 Protein Is Required for Sustained Activation of Extracellular Signal-Regulated Kinase 1/2 and Promotes Viral Virulence. J. Virol. 2012, 86, 2323–2336. [Google Scholar] [CrossRef]
- Tate, J.; Gollnick, P. The Role of Vaccinia Termination Factor and Cis-Acting Elements in Vaccinia Virus Early Gene Transcription Termination. Virology 2015, 485, 179–188. [Google Scholar] [CrossRef]
- Condit, R.C.; Moussatche, N.; Traktman, P. In a Nutshell: Structure and Assembly of the Vaccinia Virion. Adv. Virus Res. 2006, 66, 31–124. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Hruby, D.E. Vaccinia Virus A12L Protein and Its AG/A Proteolysis Play an Important Role in Viral Morphogenic Transition. Virol. J. 2007, 4, 73. [Google Scholar] [CrossRef] [PubMed]
- Ember, S.W.J.; Ren, H.; Ferguson, B.J.; Smith, G.L. Vaccinia Virus Protein C4 Inhibits NF-ΚB Activation and Promotes Virus Virulence. J. Gen. Virol. 2012, 93, 2098–2108. [Google Scholar] [CrossRef] [PubMed]
- Engelstad, M.; Smith, G.L. The Vaccinia Virus 42-KDa Envelope Protein Is Required for the Envelopment and Egress of Extracellular Virus and for Virus Virulence. Virology 1993, 194, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Gerlic, M.; Faustin, B.; Postigo, A.; Yu, E.C.-W.; Proell, M.; Gombosuren, N.; Krajewska, M.; Flynn, R.; Croft, M.; Way, M.; et al. Vaccinia Virus F1L Protein Promotes Virulence by Inhibiting Inflammasome Activation. Proc. Natl. Acad. Sci. USA 2013, 110, 7808–7813. [Google Scholar] [CrossRef]
- Morikawa, S.; Sakiyama, T.; Hasegawa, H.; Saijo, M.; Maeda, A.; Kurane, I.; Maeno, G.; Kimura, J.; Hirama, C.; Yoshida, T.; et al. An Attenuated LC16m8 Smallpox Vaccine: Analysis of Full-Genome Sequence and Induction of Immune Protection. J. Virol. 2005, 79, 11873–11891. [Google Scholar] [CrossRef]
- Jesus, D.M.; Moussatche, N.; McFadden, B.B.D.; Nielsen, C.P.; D’Costa, S.M.; Condit, R.C. Vaccinia Virus Protein A3 Is Required for the Production of Normal Immature Virions and for the Encapsidation of the Nucleocapsid Protein L4. Virology 2015, 481, 1–12. [Google Scholar] [CrossRef]
- De Silva, F.S.; Paran, N.; Moss, B. Products and Substrate/Template Usage of Vaccinia Virus DNA Primase. Virology 2009, 383, 136–141. [Google Scholar] [CrossRef]
- Van Meir, E.; Wittek, R. Fine Structure of the Vaccinia Virus Gene Encoding the Precursor of the Major Core Protein 4 A. Arch. Virol. 1988, 102, 19–27. [Google Scholar] [CrossRef]
- Blasco, R.; Sisler, J.R.; Moss, B. Dissociation of Progeny Vaccinia Virus from the Cell Membrane Is Regulated by a Viral Envelope Glycoprotein: Effect of a Point Mutation in the Lectin Homology Domain of the A34R Gene. J. Virol. 1993, 67, 3319–3325. [Google Scholar] [CrossRef]
- Brennan, G.; Kitzman, J.O.; Shendure, J.; Geballe, A.P. Experimental Evolution Identifies Vaccinia Virus Mutations in A24R and A35R That Antagonize the Protein Kinase R Pathway and Accompany Collapse of an Extragenic Gene Amplification. J. Virol. 2015, 89, 9986–9997. [Google Scholar] [CrossRef]
- Lynn, H.; Howell, L.M.; Diefenbach, R.J.; Newsome, T.P. Phototracking Vaccinia Virus Transport Reveals Dynamics of Cytoplasmic Dispersal and a Requirement for A36R and F12L for Exit from the Site of Wrapping. Viruses 2018, 10, 390. [Google Scholar] [CrossRef] [PubMed]
- Fedosyuk, S.; Grishkovskaya, I.; de Almeida Ribeiro, E.; Skern, T. Characterization and Structure of the Vaccinia Virus NF-ΚB Antagonist A46. J. Biol. Chem. 2014, 289, 3749–3762. [Google Scholar] [CrossRef]
- Rehm, K.E.; Connor, R.F.; Jones, G.J.B.; Yimbu, K.; Roper, R.L. Vaccinia Virus A35R Inhibits MHC Class II Antigen Presentation. Virology 2010, 397, 176–186. [Google Scholar] [CrossRef]
- Ryerson, M.R.; Richards, M.M.; Kvansakul, M.; Hawkins, C.J.; Shisler, J.L. Vaccinia Virus Encodes a Novel Inhibitor of Apoptosis That Associates with the Apoptosome. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Z.; Kumar, N.D.; Reggiori, F.; Khan, G. How Viruses Hijack and Modify the Secretory Transport Pathway. Cells 2021, 10, 2535. [Google Scholar] [CrossRef]
- Lin, D.; Shen, Y.; Liang, T. Oncolytic Virotherapy: Basic Principles, Recent Advances and Future Directions. Signal Transduct. Target. Ther. 2023, 8, 156. [Google Scholar] [CrossRef]
- Townsley, A.C.; Weisberg, A.S.; Wagenaar, T.R.; Moss, B. Vaccinia Virus Entry into Cells via a Low-PH-Dependent Endosomal Pathway. J. Virol. 2006, 80, 8899–8908. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Bruno, D.P.; Martens, C.A.; Porcella, S.F.; Moss, B. Simultaneous High-Resolution Analysis of Vaccinia Virus and Host Cell Transcriptomes by Deep RNA Sequencing. Proc. Natl. Acad. Sci. USA 2010, 107, 11513–11518. [Google Scholar] [CrossRef] [PubMed]
- Alkhalil, A.; Hammamieh, R.; Hardick, J.; Ichou, M.A.; Jett, M.; Ibrahim, S. Gene Expression Profiling of Monkeypox Virus-Infected Cells Reveals Novel Interfaces for Host-Virus Interactions. Virol. J. 2010, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Xuan, D.T.M.; Yeh, I.-J.; Wu, C.-C.; Su, C.-Y.; Liu, H.-L.; Chiao, C.-C.; Ku, S.-C.; Jiang, J.-Z.; Sun, Z.; Ta, H.D.K.; et al. Comparison of Transcriptomic Signatures between Monkeypox-Infected Monkey and Human Cell Lines. J. Immunol. Res. 2022, 2022, 3883822. [Google Scholar] [CrossRef]
- Tolonen, N.; Doglio, L.; Schleich, S.; Krijnse Locker, J. Vaccinia Virus DNA Replication Occurs in Endoplasmic Reticulum-Enclosed Cytoplasmic Mini-Nuclei. Mol. Biol. Cell 2001, 12, 2031–2046. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.J.; Benfield, C.T.O.; Ren, H.; Lee, V.H.; Frazer, G.L.; Strnadova, P.; Sumner, R.P.; Smith, G.L. Vaccinia Virus Protein N2 Is a Nuclear IRF3 Inhibitor That Promotes Virulence. J. Gen. Virol. 2013, 94, 2070–2081. [Google Scholar] [CrossRef]
- Teferi, W.M.; Desaulniers, M.A.; Noyce, R.S.; Shenouda, M.; Umer, B.; Evans, D.H. The Vaccinia Virus K7 Protein Promotes Histone Methylation Associated with Heterochromatin Formation. PLoS ONE 2017, 12, e0173056. [Google Scholar] [CrossRef] [PubMed]
- Soday, L.; Lu, Y.; Albarnaz, J.D.; Davies, C.T.R.; Antrobus, R.; Smith, G.L.; Weekes, M.P. Quantitative Temporal Proteomic Analysis of Vaccinia Virus Infection Reveals Regulation of Histone Deacetylases by an Interferon Antagonist. Cell Rep. 2019, 27, 1920–1933.e7. [Google Scholar] [CrossRef] [PubMed]
- Novoa, R.R.; Calderita, G.; Arranz, R.; Fontana, J.; Granzow, H.; Risco, C. Virus Factories: Associations of Cell Organelles for Viral Replication and Morphogenesis. Biol. Cell 2005, 97, 147–172. [Google Scholar] [CrossRef]
- Grimm, C.; Hillen, H.S.; Bedenk, K.; Bartuli, J.; Neyer, S.; Zhang, Q.; Hüttenhofer, A.; Erlacher, M.; Dienemann, C.; Schlosser, A.; et al. Structural Basis of Poxvirus Transcription: Vaccinia RNA Polymerase Complexes. Cell 2019, 179, 1537–1550.e19. [Google Scholar] [CrossRef]
- Ly, M.; Burgess, H.M.; Shah, S.B.; Mohr, I.; Glaunsinger, B.A. Vaccinia Virus D10 Has Broad Decapping Activity That Is Regulated by MRNA Splicing. PLoS Pathog. 2022, 18, e1010099. [Google Scholar] [CrossRef]
- Huang, T.; Nilsson, C.E.; Punga, T.; Akusjarvi, G. Functional Inactivation of the SR Family of Splicing Factors during a Vaccinia Virus Infection. EMBO Rep. 2002, 3, 1088–1093. [Google Scholar] [CrossRef] [PubMed]
