


1. Introduction
Acute ischemic stroke is a leading cause of disability and mortality, representing a major global health burden [1]. After a stroke, ischemic injury not only activates the neuroinflammation in the brain, but also results in an imbalance of the autonomic nervous system, which leads to systemic immunodepression [2,3]. Immunodepression mainly includes the transformation of T cells from pro-inflammatory T-helper (Th) 1 type to anti-inflammatory Th2 type, and the lymphocytosis in blood, spleen, and lymph nodes [4]. Immunodepression paves the way for post-stroke infections. The most common post-stroke infections are pneumonia and urinary tract infections, both reported with a percentage of 10% [5].
Stroke-associated pneumonia (SAP) is the most notable post-stroke infection for its significant association with poor stroke outcome [6,7,8]. SAP could increase stroke mortality and the length of in-hospital stay, affecting self-care ability [9,10]. Based on the known risk factors for SAP, such as age, dysphagia, and stroke severity [11,12], prediction models have been developed to identify patients at an increased risk for SAP, such as the Acute Ischemic Stroke-Associated Pneumonia Score [13] (AIS-APS), and the Age, Atrial fibrillation, Dysphagia, Sex, Stroke Severity Score [14] (A2DS2). Previous studies have proved the validity of AIS-APS and A2DS2 in predicting SAP [15,16]. In a recent meta-analysis, AIS-APS and A2DS2 showed moderate predictive accuracy for SAP [17]. To improve the discriminatory ability of the present models, more predictive risk factors of SAP need to be included.
Recent studies have demonstrated that a stroke could change the composition of patients’ gut microbiota, with an increase in opportunistic pathogens and a decrease in anti-inflammatory bacteria [18,19]. Furthermore, a stroke could reduce the transepithelial resistance of the gut barrier, disturb the immune homeostasis of the intestine, and promote the enrichment of proinflammatory bacteria [20]. The gut dysbiosis enhances the systemic inflammation in turn and induces the post-stroke infection [21]. Stanley et al. reported the activation of the sympathetic nervous system increased intestinal permeability after a stroke, leading to the translocation of the microbiota from the intestinal tract to the lung, ultimately resulting in post-stroke pneumonia [22]. The gut microbiota are thought to play a critical role in the occurrence and development of SAP, and several studies have proved it [23,24]. The feature of gut microbiota in stroke patients may be an essential predictor of SAP.
Therefore, this study aimed to (1) compare the composition of gut microbiota in acute ischemic stroke patients with and without SAP, (2) identify the patterns of altered gut microbiota in SAP patients, and (3) discern the improvement of predictive ability by integrating specific gut microbiota for SAP into conventional risk prediction models (i.e., AIS-APS and A2DS2).
2. Methods
2.1. Study Participants
This prospective observational cohort study was conducted in Nanjing First Hospital from May 2018 to June 2019. The inclusion criteria were as follows: (1) aged 50 years or older; (2) admitted within 72 h of symptom onset with a magnetic resonance imaging-confirmed diagnosis of acute anterior ischemic stroke; (3) have lived in Nanjing for at least six months. The exclusion criteria included: (1) use of antibiotics, probiotics, corticosteroids, or immunosuppressants in the last 30 days before admission; (2) a history of immune diseases, severe liver and kidney failure, or malignant tumors; (3) acute hemorrhagic stroke; (4) unavailable blood or stool samples. This study was approved by the Ethical Review Board of Nanjing First Hospital (Nanjing, China). The written informed consent was provided by all patients (or their immediate family members) before participating in this study.
SAP was diagnosed within seven days after stroke onset by two neurologists according to the 2015 Consensus Group criteria [25], with evidence of sputum culture or radiological signs of pulmonary infection on the chest-computed tomography. Participants who had SAP were included in the SAP group, while those without SAP were in the non-SAP (NSAP) group.
2.2. Data Collection
The basic information and medical histories were obtained from participants or their immediate family members through face-to-face conversations. All serum and stool samples were collected within 24 h of admission before the treatment with antibiotics or probiotics. The blood parameters were measured in the central laboratory of Nanjing First Hospital by the laboratory technicians who were blind to the clinical information. The hematological parameters including of white blood counts (WBCs), neutrophil, and lymphocyte counts were analyzed by Hematology Analyzer (BC-6900, Mindray, Shenzhen, China). The blood glucose and CRP were assayed by Fully Automated Biochemistry Analyzer (C16000, Abbott, Abbott Park, IL, USA).
The clinical subtype of stroke was determined by the Oxfordshire Community Stroke Project (OCSP) classification [26]. The etiology of stroke was described using the Trial of Org 10172 in Acute Stroke Treatment (TOAST) classification [27]. The neurological function was evaluated by modified Rankin Scale (mRS). The severity of stroke was assessed by the National Institute of Health Stroke Scale score (NIHSS). The level of consciousness was assessed by Glasgow Coma Scale (GCS), a 15-point scale composed of eye, verbal, and motor responses [28]. AIS-APS and A2DS2 were used to predict the risk of SAP. AIS-APS is a 34-point score based on demographics, medical history, pre-stroke mRS, stroke features, and admission glucose level [13]. A2DS2 score is a 10-point scale calculated as follows [14]: age ≥ 75 years = 1, atrial fibrillation = 1, dysphagia = 2, male sex = 1, NIHSS score of 0–4 = 0, NIHSS score of 5–15 = 3, and NIHSS ≥ 16 = 5. All clinical scores were evaluated by two experienced neurologists.
2.3. DNA Extraction and Sequencing
All 135 fecal samples underwent DNA extraction and sequencing in July 2019. Using the QIAamp® DNA Stool Mini Kit (Qiagen, Hilden, Germany), the fecal DNA was extracted from stool samples according to the instructions. Briefly, the stool samples were lysed in argininosuccinate lyase buffer, and InhibitEX was used to adsorb impurities. Next, protease K was used to digest proteins, and DNA was purified with a two-step wash. After centrifugation, DNA was finally eluted from the spin column in the low-salt buffer. The concentration of DNA was calculated by measuring the absorbance of DNA eluate at 260 nm through the Nanodrop. The integrity was verified by 0.8% agarose gel electrophoresis.
The V3 to V4 hypervariable regions of the bacterial 16S rRNA gene were amplified by polymerase chain reaction (PCR) using the forward primer (5′-CCTACGGGNGGCWGCAG-3′) and the reverse primer (5′-GACTACHVGGGTATCTAATCC-3′) [29]. The amplified products were detected by gel electrophoresis and purified by the Agencourt AMPure XP Kit (Beckman Coulter, Brea, CA, USA). The index of purified products was performed in the 16S V3-V4 library. The [email protected] Fluorometer (Thermo Scientific, Waltham, MA, USA) and Agilent Bioanalyzer 2100 systems (USA) were used to evaluate the library quality. Using the 2 × 250 bp paired-end read protocol, high throughput sequencing was performed on the Illumina Miseq platform. This work was supported by the Shanghai Genesky Biotechnology Company (Shanghai, China).
2.4. Bioinformatics and Statistical Analyses
After quality filtering merging, UPARSE was used to cluster the raw reads into operational taxonomic units (OTUs) with a 97% similarity. All OTUs were classified by Mothur, according to Ribosomal Database Project (RDP) Release 9. Then, the Alpha diversities, including Chao, ACE, Shannon, Simpson, and Coverage index, were analyzed using Mothur. The Beta diversities were analyzed by permutational multivariate analysis of variance (PERMANOVA) and presented visually by principal coordinate analysis (PCoA). The Metastats and Linear discriminant analysis (LDA) Effect Size (LEfSe) was used to determine the microbial features between the two groups. The absolute values of logarithmic LDA score > 2 and p-value < 0.05 were considered statistically significant. The p values were adjusted with the Benjamini–Hochberg false discovery rate (FDR) correction for multiple testing. These analyses were performed on R version 3.4.3 (Vegan package).
The student t-test or Mann–Whitney U test was used to compare the statistical differences between the two groups for continuous variables. Fisher’s exact probability test or chi-squared test was applied for categorical variables. Also, Spearman’s rank correlation coefficient analysis was used to measure the correlation between gut microbiota and the prediction scores. The receiver operating characteristic curve (ROC) was performed to assess the predictive performance of gut microbiota. The Net Reclassification Improvement (NRI) and Integrated Discrimination Improvement (IDI) indices were generated to evaluate the improvement of predictive abilities after adding specific gut microbiota into AIS-APS and A2DS2. A p-value of <0.05 was considered to be statistically significant. All the statistical analyses were performed on SPSS 22.0 for Windows (IBM Inc., New York, NY, USA).
3. Results
3.1. Baseline Characteristics
We screened 732 patients with acute ischemic stroke and recruited 135 patients from May 2018 to June 2019. A total of 43 patients developed SAP (31.8%). Univariate analyses showed that patients in the SAP group were older than those in the NSAP group (71.5 vs. 66.3 years, p = 0.006). The SAP group, in comparison to the NSAP group, had a smaller percentage of male patients (51.2% vs. 75.0%, p = 0.006), and were more likely to have atrial fibrillation (44.2% vs. 15.2%, p < 0.001), dysphagia (39.5% vs. 8.7%, p < 0.001), and speech disorders (86.0% vs. 66.3%, p = 0.017). Patients with SAP had higher levels of WBC (9.2 vs. 7.4 × 109/L, p = 0.001), neutrophil counts (5.6 vs. 4.4 × 109/L, p < 0.001), C-reactive protein (9.4 vs. 2.5 ug/mL, p < 0.001), and neutrophil-to-lymphocyte ratios (NLRs) (4.8 vs. 2.7, p < 0.001) compared to the NSAP patients. Furthermore, higher NIHSS scores (11.0 vs. 3.0, p < 0.001) and lower GCS scores (8–15 vs. 15–15, p < 0.001) on admission, higher AIS-APS scores (11.0 vs. 5.0, p < 0.001), and higher A2DS2 scores (5.0 vs. 1.0, p < 0.001) were found in the SAP group compared with the NSAP group .
3.2. Altered Gut Microbiota in the SAP Patients
The α-diversity indices of gut microbiota showed no significant group differences (Figure 1), except for the Simpson index, which indicated a lower diversity of the microbial community in the SAP group than that of the NSAP group (p = 0.022). The results of PCoA revealed that Bray distances (Coefficient of determination (R2) = 0.014, p = 0.007), Jaccard distances (R2 = 0.010, p = 0.035), and weighted uniFrac distances (R2 = 0.050, p < 0.001) were significantly different between the two groups.
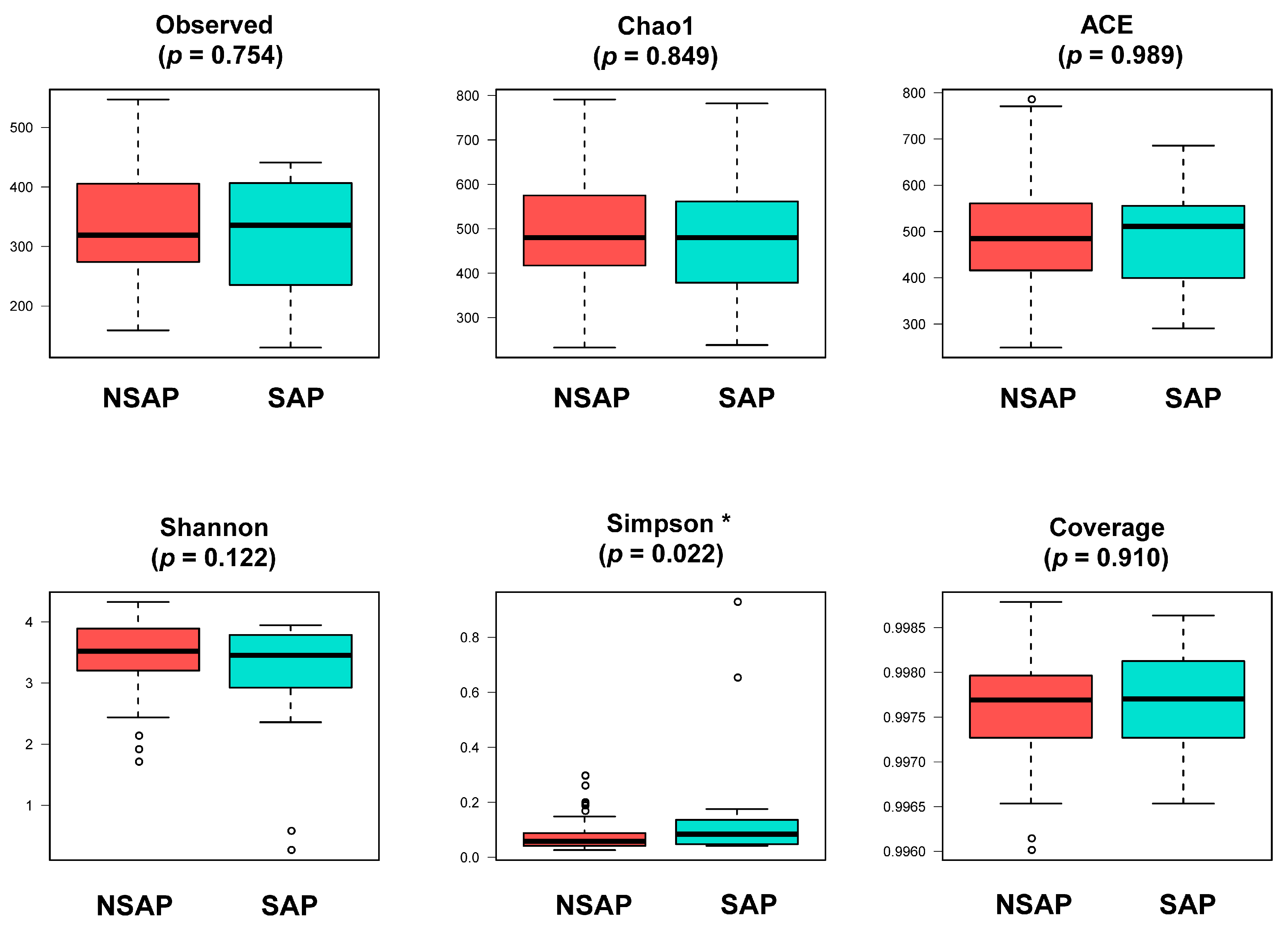
Figure 1. Comparison of α-diversity of gut microbiota between two groups. Abbreviation: SAP, stroke-associated pneumonia group; NSAP, non-SAP group. Note: * p < 0.05.
There were 112 unique OTUs in the SAP group, 979 unique OTUs in the NSAP group, and 1511 shared OTUs in both groups, as shown in the Venn diagram . The phyla of Firmicutes, Bacteroidetes, and Proteobacteria comprised most of the gut microbiota community. The Metastats analysis showed that the relative abundance of phylum Bacteroidetes in the NSAP group was higher than in the SAP group (p = 0.020) . At the family level, the abundances of Bacteroidaceae (p = 0.007), Veillonellaceae (p = 0.029), and Sutterellaceae (p = 0.014) were relatively lower in the SAP group . At the genus level, Bacteroides (p = 0.006), Coprococcus (p = 0.032), Fusicatenibacter (p = 0.039), Butyricicoccus (p < 0.001), Butyricimonas (p = 0.046), and Clostridium_XlVb (p = 0.018) were more enriched in the NSAP group (Figure 2). Also, the results of LEfSe analysis and LDA score showed that the relative abundances of class Bacilli, order Lactobacillales, family Corynebacteriaceae, genus Corynebacterium, and the species Clostridium innocuum in the SAP group were higher than those in the NSAP group (Figure 3).
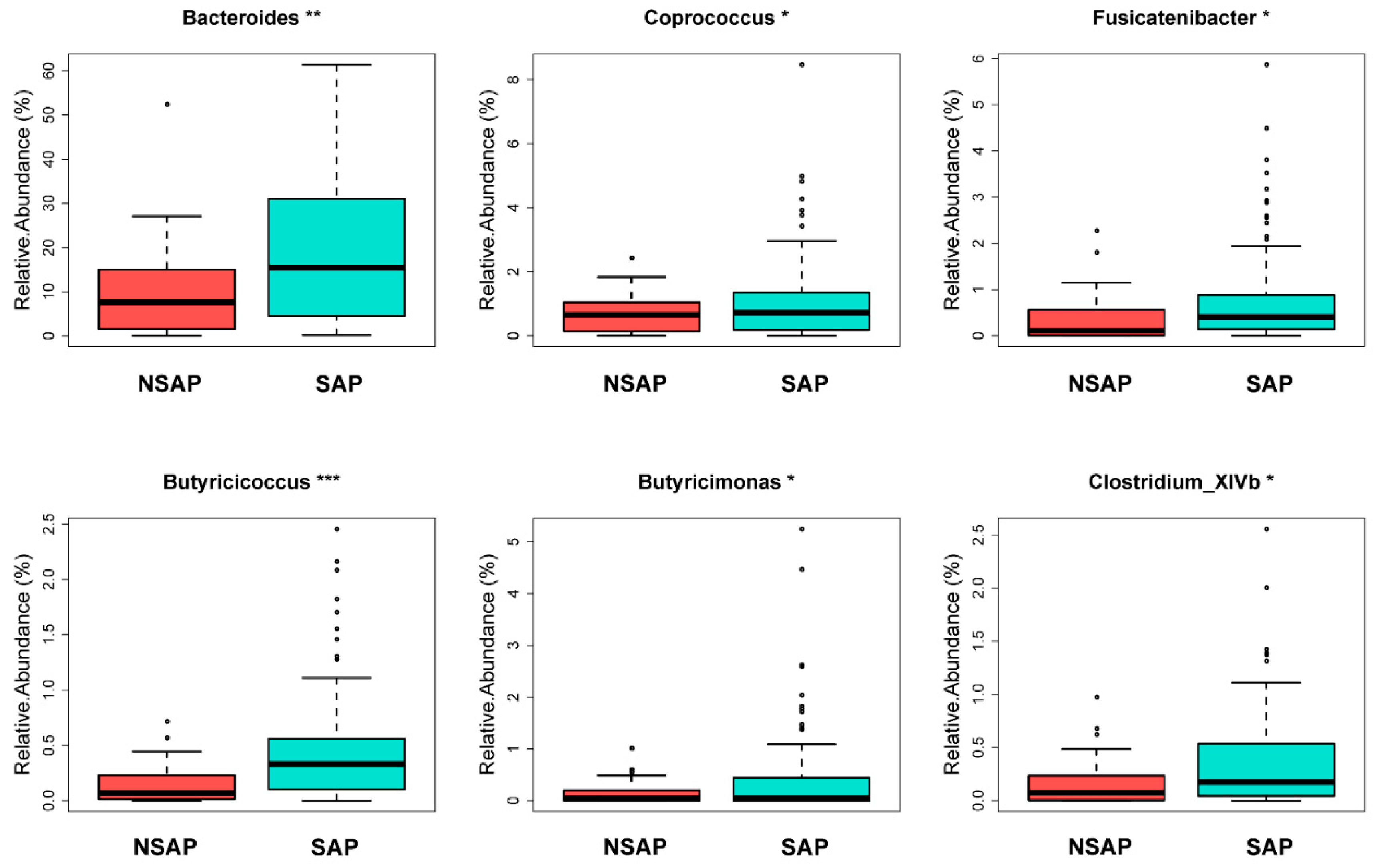
Figure 2. Gut microbiota with significantly different relative abundance between the two groups at genus level in Metastats analysis. Abbreviation: SAP, stroke-associated pneumonia; NSAP, non-SAP group. Note: * p < 0.05, ** p < 0.01, *** p < 0.001.
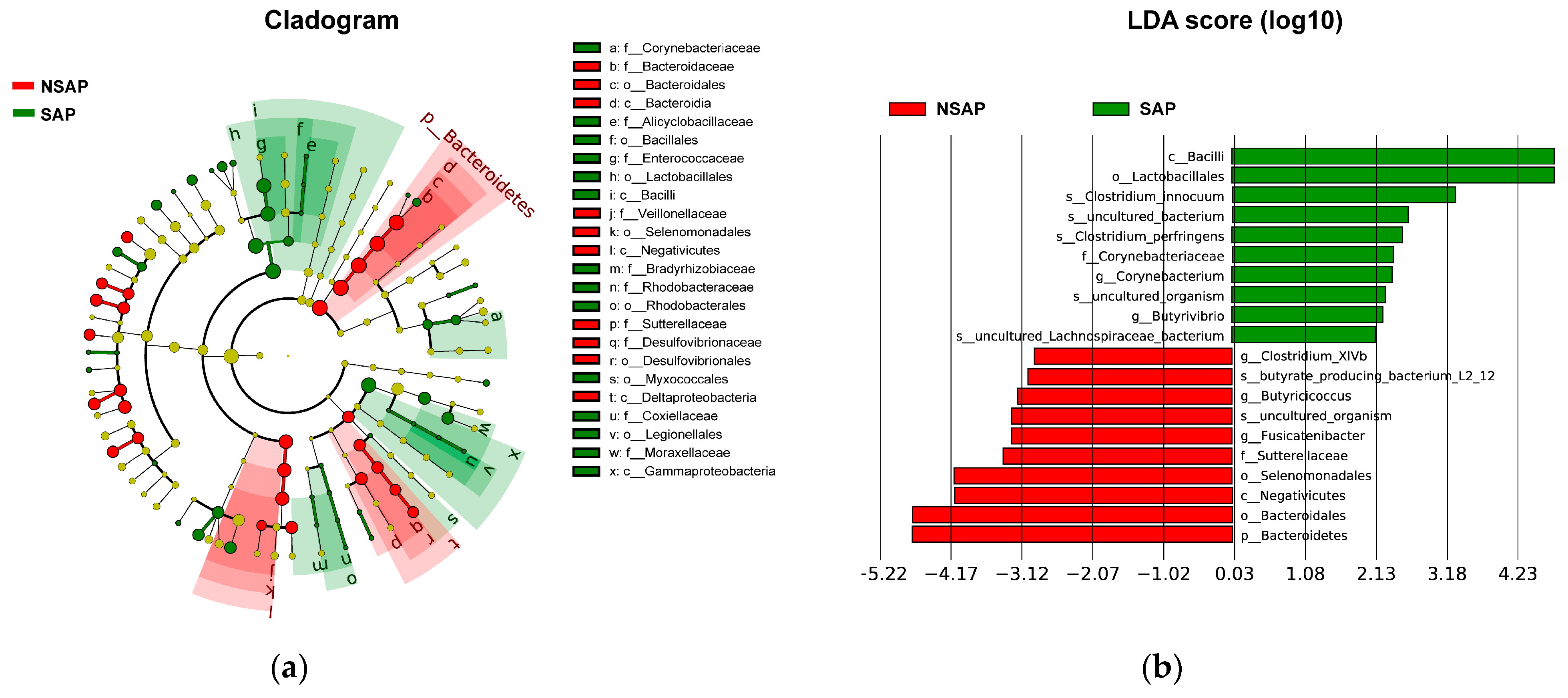
Figure 3. LEfSe of gut microbiota with significant difference between two groups. (a). Cladogram of differential bacteria; (b). Significantly differential bacteria with LDA scores > 2 between two groups. Abbreviation: LDA, Linear discriminant analysis; LEfSe, LDA Effect Size; SAP, stroke-associated pneumonia; NSAP, non-SAP group.
3.3. Gut Microbiota Correlated with SAP Predictive Scores
Both scores of AIS-APS and A2DS2 were higher in the SAP group. The analyses on their correlation with the 50 most common genera showed that the genus Bacteroides was negatively correlated with AIS-APS (r = −0.197, p = 0.02) and A2DS2 (r = −0.320, p < 0.001). The same results were obtained for genus Fusicatenibacter (r = −0.25, p = 0.003 in AIP-APS; r = −0.22, p = 0.010 in A2DS2), Butyricicoccus (r = −0.24, p = 0.004 in AIP-APS; r = −0.219, p = 0.010 in A2DS2), and Clostridium_XlVb (r = −0.230, p = 0.007 in AIP-APS; r = −0.254, p = 0.003 in A2DS2). The genus Enterococcus had a significant positive correlation with both AIS-APS and A2DS2 scores (r = 0.297, p < 0.001 in AIS-APS; r = 0.360, p < 0.001 in A2DS2), though it did not show any significant difference between the SAP and NSAP groups (Figure 4).
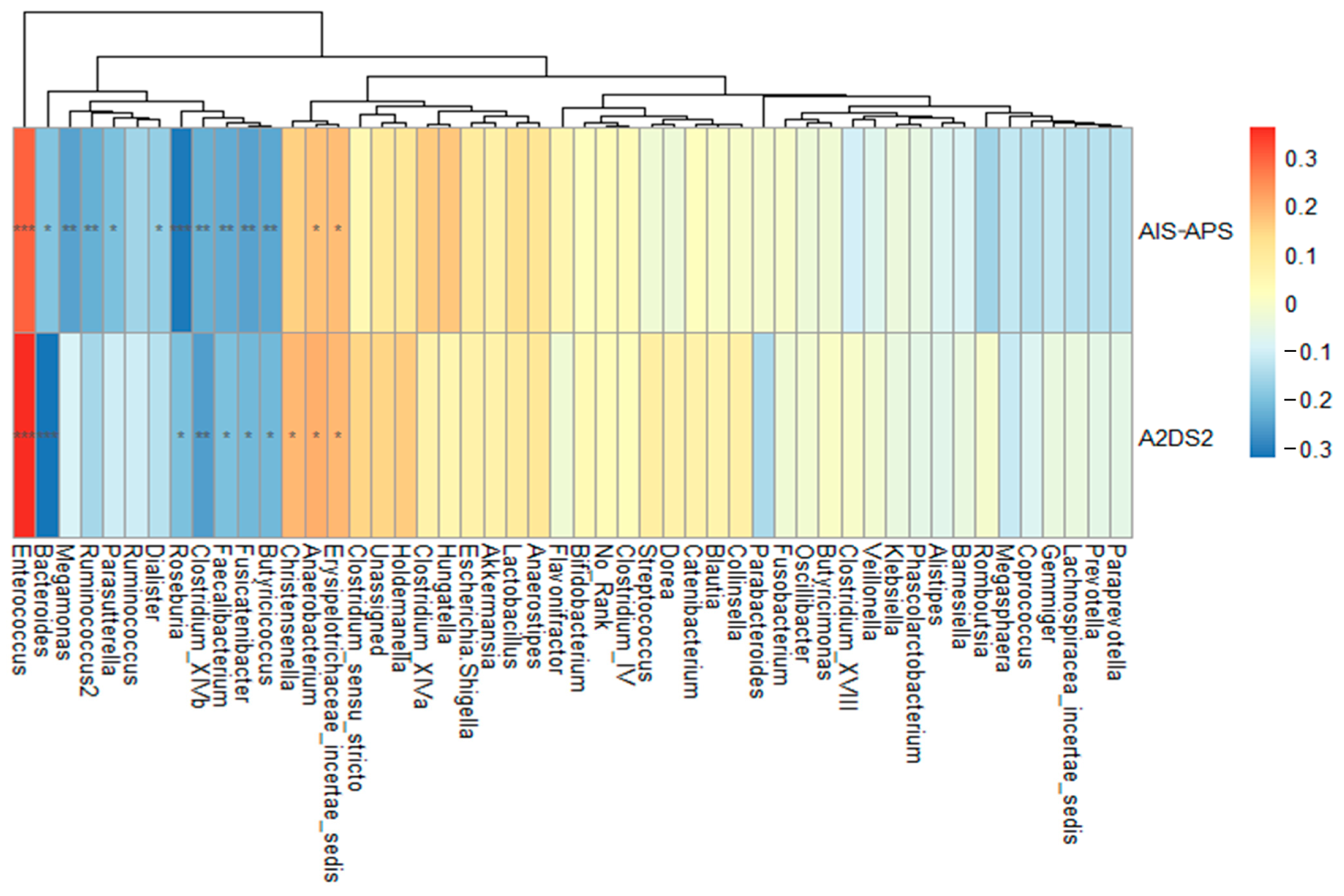
Figure 4. Spearman’s rank correlation analysis between the most 50 common genera and two predictive scores. Abbreviation: AIS-APS, the Acute Ischemic Stroke-Associated Pneumonia Score; A2DS2, the Age, Atrial fibrillation, Dysphagia, Sex, Stroke Severity Score. Note: * p < 0.05; ** p < 0.01; *** p < 0.001.
3.4. Predictive Performance of Gut Microbiota for SAP
We tested the predictive validity of the six genera with significant group differences in Metastats analysis, including Bacteroides, Coprococcus, Fusicatenibacter, Butyricicoccus, Butyricimonas, and Clostridium-IVb. The area under the curve (AUC) of the differential genera was 0.699 (Standard Error (SE): 0.046, 95% Confidence Interval (CI): 0.609–0.790), while the AUCs of AIS-APS (SE: 0.047, 95% CI: 0.684–0.870) and A2DS2 (SE: 0.050, 95% CI: 0.679–0.874) were both 0.777. After adding the differential genera into AIS-APS, the AUC of the integrated model was improved to 0.813 (SE: 0.040, 95% CI: 0.735–0.891, as seen in Figure 5a). The results of IDI showed that the predictive ability of the model was significantly improved (IDI = 0.038, 95% CI: 0.006–0.070, p = 0.018), while NRI showed a borderline significant improvement (Continuous NRI = 0.333, 95% CI: −0.003–0.700, p = 0.052, . The ability of A2DS2 to predict SAP was also significantly improved after adding the differential genera, with the AUC improved to 0.816 (SE: 0.041, 95% CI: 0.735–0.897, as seen in Figure 5b), NRI 57.5% (95% CI: 0.245–0.906, p < 0.001), and IDI 4.3% (95% CI: 0.012–0.075, p = 0.007, as seen in .
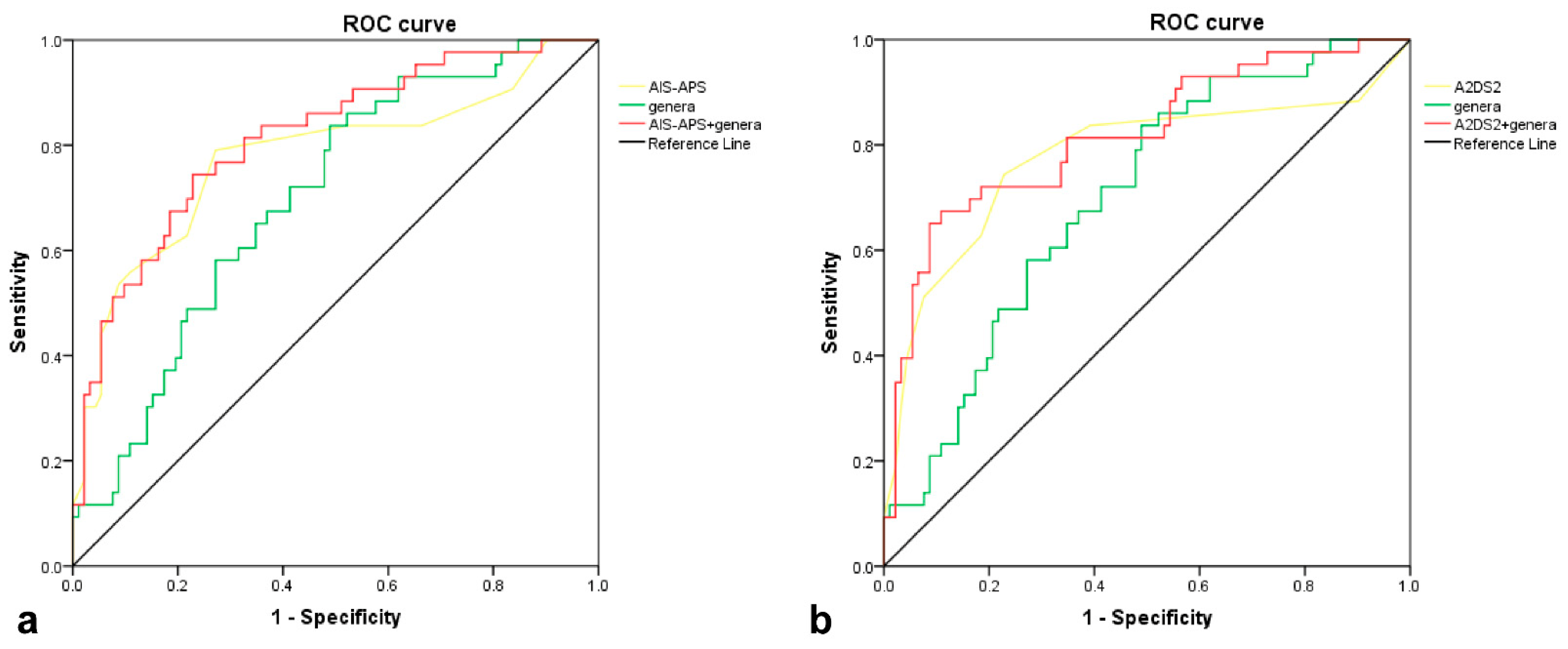
Figure 5. ROC curves of differential genera, 2 predictive scores, and the combination of genera and scores in predicting SAP. (a). ROC of genera (AUC = 0.699), AIS-APS (AUC = 0.777), and the combination of genera and AIS-APS (AUC = 0.813); (b). ROC of genera (AUC = 0.699), A2DS2 (AUC = 0.777), and the combination of genera and A2DS2 (AUC = 0.816). Genera consist of 6 bacteria at genus level, Bacteroides, Coprococcus, Fusicatenibacter, Butyricicoccus, Butyricimonas, and Clostridium-IVb, with significant difference between two groups in Metastats analysis. Abbreviation: ROC, receiver operating characteristic curve; AIS-APS, the Acute Ischemic Stroke-Associated Pneumonia Score; A2DS2, the Age, Atrial fibrillation, Dysphagia, Sex, Stroke Severity Score; SAP, stroke-associated pneumonia.
4. Discussion
In this prospective observational cohort study, we explored the baseline differences in gut microbiota between SAP and NSAP patients. The two groups had significant differences in α-diversity and β-diversity, showing lower bacteria richness in SAP patients. Specifically, the genera Bacteroides, Coprococcus, Fusicatenibacter, Butyricicoccus, Butyricimonas, and Clostridium_XlVb were less abundant in the SAP group, while the abundances of family Corynebacteriaceae, genus Corynebacterium, and species Clostridium_innocuum were higher in the SAP group. Furthermore, the six decreased genera in the SAP group compared to the NSAP group could improve the predictive ability of AIS-APS and A2DS2.
Previous studies have revealed an alteration in gut microbiota composition after a stroke, which can differ depending on the severity of the stroke [18,19,30]. As a stress event, the stroke will activate the sympathetic nervous system, hypothalamic–pituitary–adrenal axis, and enteric nervous system, increasing the permeability of the intestinal mucosal barrier and the translocation of gut microbiota [20,31]. With the close association between the gut and the lung, gut dysbiosis could influence pulmonary health [32]. Chen et al. reported that commensal gut microbiota played a vital role in immune defense against Escherichia Coli pneumonia by inducing the expression of Toll-like receptor 4 (TLR) and activation of nuclear factor κB [33]. By stimulating TLR, gut microbiota can regulate the immune response of respiratory mucosa against influenza virus infection [34]. As one of the most significant and abundant commensal flora in the human intestine, the phyla Bacteroidetes can stimulate the TLR signaling pathways and regulate Treg cells [35,36]. Our study detected the early changes in gut microbiota in SAP patients, particularly the decrease in phyla Bacteroidetes, which suggested impaired immune surveillance preceding SAP.
The genera Bacteroides, Coprococcus, Butyricicoccus, Butyricimonas, and Fusicatenibacter, of which the abundances were lower in SAP patients, are all SCFAs-producing bacteria [37,38,39,40]. Among them, Fusicatenibacter and Butyricicoccus were negatively correlated with AIS-APS and A2DS2. A similar decrease in SCFAs-producing bacteria has been detected in patients with ischemic stroke [18] and post-stroke infection [24]. SCFAs are essential for their anti-inflammatory properties and immunomodulatory effects, such as suppressing the production of pro-inflammatory Interleukin-6 (IL-6) and inducing the production of anti-inflammatory IL-10 [41,42,43,44]. For instance, as one of the SCFAs, acetate was reported to activate the interferon-β response to enhance the antiviral effect on pulmonary epithelial cells [45]. While butyrate, another member of SCFAs, was positively associated with a decreased risk of lower respiratory tract infection due to its anti-inflammatory properties [46]. The transplantation of SCFAs-producing bacteria could improve neurological dysfunction and reduce inflammation in aged mice with middle cerebral artery occlusion [47]. Furthermore, increasing the levels of SCFAs through symbiotic therapy could reduce the incidence of enteritis and ventilator-associated pneumonia in sepsis patients [48]. Counterintuitively, several well-known SCFAs-producing bacteria like Roseburia were not differential between the two groups in our study, which was inconsistent with a recent study [23]. We thought that this may be attributable to the discrepancy between two cohorts, such as stroke severity and dietary structures, which could influence gut microbiota.
The alteration in the gut microbiota after a stroke, which involves the growth of pro-inflammatory bacteria, could affect immune homeostasis [49]. Our study found that several pathogenic bacteria were enriched in the SAP group, including the family Corynebacteriaceae, the genus Corynebacterium, and the species Clostridium innocuum. The genus Corynebacterium, which belongs to the family Corynebacteriaceae, is a potential pathogen due to its ability to produce the diphtheria toxin [50]. Clostridium Innocuum was reported to be cytotoxic by triggering cell death through apoptosis, leading to a number of infectious diseases, such as intra-abdominal infection, pylephlebitis, and empyema [51]. Highet et al. described that Clostridium Innocuum could increase the susceptibility to infection [52]. Recently, the infection of Clostridium Innocuum was reported to be associated with severe gastrointestinal complications and extraintestinal infections [53]. The accumulation of pro-inflammatory bacteria in stroke patients suggested that the imbalance in the immunologic barrier [54] may be a cause of SAP.
Compared to NSAP patients, SAP patients scored higher on two conventional predictive scores of AIS-APS and A2DS2. These scores were negatively correlated with the SCFAs-producing bacteria Bacteroides, Fusicatenibacter, and Butyricicoccus. Moreover, the six decreased genera in the SAP group, Bacteroides, Coprococcus, Fusicatenibacter, Butyricicoccus, Butyricimonas, and Clostridium-IVb, were mostly SCFA-producing bacteria (except Clostridium-IVb, which contains both beneficial and pathogenic species [55]). Haak et al. established that the decline in SCFAs-producing bacteria was an independent predictor of post-stroke infection [24]. Xia et al. showed that decreased SCFAs-producing genus Roseburia and increased pathogenic bacteria were correlated with the risk of SAP [23]. Our study confirmed their observations and further proved the predictive value of the above bacteria when integrated into the predictive scores. When the group-differential genera were incorporated, the predictive performances of AIS-APS and A2DS2 were both enhanced. This would imply that an altered gut microbiota, especially a lower abundance of SCFA-producing bacteria, could be a predictor of SAP. The existing predictive scores mainly focus on the clinical symptoms and may ignore the change in homeostasis prior to SAP onset [17]. Incorporating altered gut microbiota at baseline may pioneer new ways of early predictive models of SAP after further clinical verification.
Interestingly, we found that the relative abundance of the genus Enterococcus was significantly correlated with AIS-APS and A2DS2. Enterococcus was thought to be one of the opportunistic commensal bacteria associated with several infections like urinary tract infection, endocarditis, and biliary tract infection [56]. Enterococcus was also reported to be enriched in pneumonia patients after hypertensive intracerebral hemorrhage [57] and ischemic stroke [23], possibly due to the immunodepression after the stroke [58]. However, Enterococcus was not significantly different between SAP and NSAP groups in our cohort. This may be attributable to the regional differences between participants from previous studies, whose different diet structures may have affected the gut microbiota composition.
Our single-center study had several limitations. Firstly, we did not characterize the alterations in stool pH, inflammatory markers in stool, leaky gut markers, and metabolites such as SCFAs in the fecal samples. Secondly, diet has been demonstrated to be an essential factor influencing the composition of the gut microbiota [59]. Although we enrolled residents with presumably consistent dietary structures, the dietary intake data of each participant were not collected. Thirdly, we collected fecal samples at a single time point, and the dynamic changes in the gut microbiota were unavailable due to the limited funds. Fourthly, our study was an observational analysis without functional data to show causal links; hence, the results should be treated with caution until the precise mechanism of brain–gut–lung communication is revealed. Finally, due to the small sample size of this study, the reliability of the early prediction and diagnosis data needs to be further verified.
5. Conclusions
In conclusion, the baseline abundance of SCFAs-producing gut microbiota was decreased in patients who developed SAP within a week after the onset of acute ischemic stroke. The differential gut microbiota between SAP and NSAP added prognostic value to the conventional SAP risk scores of AIS-APS and A2DS2. This study emphasized the role of the altered post-stroke gut microbiota in predicting SAP, which merits attention in further clinical research.
References
- Feigin, V.L.; Norrving, B.; Mensah, G.A. Global Burden of Stroke. Circ. Res. 2017, 120, 439–448. [Google Scholar] [CrossRef]
- Westendorp, W.F.; Dames, C.; Nederkoorn, P.J.; Meisel, A. Immunodepression, Infections, and Functional Outcome in Ischemic Stroke. Stroke 2022, 53, 1438–1448. [Google Scholar] [CrossRef] [PubMed]
- Faura, J.; Bustamante, A.; Miró-Mur, F.; Montaner, J. Stroke-induced immunosuppression: Implications for the prevention and prediction of post-stroke infections. J. Neuroinflamm. 2021, 18, 127. [Google Scholar] [CrossRef]
- Prass, K.; Meisel, C.; Höflich, C.; Braun, J.; Halle, E.; Wolf, T.; Ruscher, K.; Victorov, I.V.; Priller, J.; Dirnagl, U.; et al. Stroke-induced immunodeficiency promotes spontaneous bacterial infections and is mediated by sympathetic activation reversal by poststroke T helper cell type 1-like immunostimulation. J. Exp. Med. 2003, 198, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Westendorp, W.F.; Nederkoorn, P.J.; Vermeij, J.D.; Dijkgraaf, M.G.; van de Beek, D. Post-stroke infection: A systematic review and meta-analysis. BMC Neurol. 2011, 11, 110. [Google Scholar] [CrossRef] [PubMed]
- Vermeij, F.H.; Scholte op Reimer, W.J.; de Man, P.; van Oostenbrugge, R.J.; Franke, C.L.; de Jong, G.; de Kort, P.L.; Dippel, D.W.; Netherlands Stroke Survey, I. Stroke-associated infection is an independent risk factor for poor outcome after acute ischemic stroke: Data from the Netherlands Stroke Survey. Cerebrovasc. Dis. 2009, 27, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.; Hand, P. Infection after acute stroke is associated with poor short-term outcome. Acta Neurol. Scand. 2007, 115, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Hilker, R.; Poetter, C.; Findeisen, N.; Sobesky, J.; Jacobs, A.; Neveling, M.; Heiss, W.D. Nosocomial pneumonia after acute stroke: Implications for neurological intensive care medicine. Stroke 2003, 34, 975–981. [Google Scholar] [CrossRef]
- Katzan, I.L.; Cebul, R.D.; Husak, S.H.; Dawson, N.V.; Baker, D.W.J.N. The effect of pneumonia on mortality among patients hospitalized for acute stroke. Neurology 2003, 60, 620. [Google Scholar] [CrossRef] [PubMed]
- Finlayson, O.; Kapral, M.; Hall, R.; Sllani, E.A.; Selchen, D.; Saposnik, G.J.N. Risk factors, inpatient care, and outcomes of pneumonia after ischemic stroke. Neurology 2011, 77, 1338. [Google Scholar] [CrossRef]
- André, K.; Lev, M.H.; Seyedmehdi, P.; Betensky, R.A.; Jing, Q.; Shihab, M.; Schwamm, L.H.; Jens, M.J.P.O. Hospital Acquired Pneumonia Is Linked to Right Hemispheric Peri-Insular Stroke. PLoS ONE 2013, 8, e71141. [Google Scholar]
- Chumbler, N.R.; Williams, L.S.; Wells, C.K.; Lo, A.; Nadeau, S.; Peixoto, A.J.; Gorman, M.; Boice, J.L.; Concato, J.; Bravata, D.M.J.N. Derivation and Validation of a Clinical System for Predicting Pneumonia in Acute Stroke. Neuroepidemiology 2010, 34, 193–199. [Google Scholar] [CrossRef]
- Ji, R.; Shen, H.; Pan, Y.; Wang, P.; Liu, G.; Wang, Y.; Li, H.; Wang, Y.; China National Stroke Registry, I. Novel risk score to predict pneumonia after acute ischemic stroke. Stroke 2013, 44, 1303–1309. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, S.; Malzahn, U.; Harms, H.; Koennecke, H.C.; Berger, K.; Kalic, M.; Walter, G.; Meisel, A.; Heuschmann, P.U.; Berlin Stroke, R.; et al. Development of a clinical score (A2DS2) to predict pneumonia in acute ischemic stroke. Stroke 2012, 43, 2617–2623. [Google Scholar] [CrossRef]
- Helmy, T.A.; Abd-Elhady, A.E.; Abdou, M.J.J.o.S.; Association, C.D.t.O.J.o.N.S. Prediction of Ischemic Stroke-Associated Pneumonia: A Comparison between 3 Scores. J. Stroke Cerebrovasc. Dis. 2016, 25, 2756–2761. [Google Scholar] [CrossRef]
- Zapata-Arriaza, E.; Moniche, F.; Blanca, P.G.; Bustamante, A.; Escudero-Martinez, I.; Ucles, O.; Ollero-Ortiz, A.; Sanchez-Garcia, J.A.; Gamero, M.A.; Quesada, A.; et al. External Validation of the ISAN, A2DS2, and AIS-APS Scores for Predicting Stroke-Associated Pneumonia. J. Stroke Cerebrovasc. Dis. 2018, 27, 673–676. [Google Scholar] [CrossRef]
- Ni, J.; Shou, W.; Wu, X.; Sun, J. Prediction of stroke-associated pneumonia by the A2DS2, AIS-APS, and ISAN scores: A systematic review and meta-analysis. Expert. Rev. Respir. Med. 2021, 15, 1461–1472. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Liao, S.X.; He, Y.; Wang, S.; Xia, G.H.; Liu, F.T.; Zhu, J.J.; You, C.; Chen, Q.; Zhou, L.; et al. Dysbiosis of Gut Microbiota With Reduced Trimethylamine-N-Oxide Level in Patients With Large-Artery Atherosclerotic Stroke or Transient Ischemic Attack. J. Am. Heart Assoc. 2015, 4, e002699. [Google Scholar] [CrossRef]
- Yamashiro, K.; Tanaka, R.; Urabe, T.; Ueno, Y.; Yamashiro, Y.; Nomoto, K.; Takahashi, T.; Tsuji, H.; Asahara, T.; Hattori, N. Gut dysbiosis is associated with metabolism and systemic inflammation in patients with ischemic stroke. PLoS ONE 2017, 12, e0171521. [Google Scholar] [CrossRef]
- Diaz-Marugan, L.; Gallizioli, M.; Marquez-Kisinousky, L.; Arboleya, S.; Mastrangelo, A.; Ruiz-Jaen, F.; Pedragosa, J.; Casals, C.; Morales, F.J.; Ramos-Romero, S.; et al. Poststroke Lung Infection by Opportunistic Commensal Bacteria Is Not Mediated by Their Expansion in the Gut Microbiota. Stroke 2023, 54, 1875–1887. [Google Scholar] [CrossRef]
- Xu, K.; Gao, X.; Xia, G.; Chen, M.; Yin, J.J.G. Rapid gut dysbiosis induced by stroke exacerbates brain infarction in turn. Gut 2021, 70, 1486–1494. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Mason, L.J.; Mackin, K.E.; Srikhanta, Y.N.; Lyras, D.; Prakash, M.D.; Nurgali, K.; Venegas, A.; Hill, M.D.; Moore, R.J.; et al. Translocation and dissemination of commensal bacteria in post-stroke infection. Nat. Med. 2016, 22, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.H.; Zhang, M.S.; Wu, Q.H.; Wang, H.D.; Zhou, H.W.; He, Y.; Yin, J. Dysbiosis of Gut Microbiota Is an Independent Risk Factor of Stroke-Associated Pneumonia: A Chinese Pilot Study. Front. Cell Infect. Microbiol. 2021, 11, 715475. [Google Scholar] [CrossRef] [PubMed]
- Haak, B.W.; Westendorp, W.F.; van Engelen, T.S.R.; Brands, X.; Brouwer, M.C.; Vermeij, J.D.; Hugenholtz, F.; Verhoeven, A.; Derks, R.J.; Giera, M.; et al. Disruptions of Anaerobic Gut Bacteria Are Associated with Stroke and Post-stroke Infection: A Prospective Case-Control Study. Transl. Stroke Res. 2021, 12, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Kishore, A.K.; Vail, A.; Chamorro, A.; Garau, J.; Hopkins, S.J.; Di Napoli, M.; Kalra, L.; Langhorne, P.; Montaner, J.; et al. Diagnosis of Stroke-Associated Pneumonia: Recommendations From the Pneumonia in Stroke Consensus Group. Stroke 2015, 46, 2335–2340. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, J.B.C.; Mohr, J.P.; Timbó, F.B.; Nepomuceno, C.R.; Moreira, J.; Timbó, I.; Lima, F.O.; Silva, G.S.; Bamford, J. Oxfordshire Community Stroke Project Classification: A proposed automated algorithm. Eur. Stroke J. 2021, 6, 160–167. [Google Scholar] [CrossRef]
- Adams, H.P., Jr.; Bendixen, B.H.; Kappelle, L.J.; Biller, J.; Love, B.B.; Gordon, D.L.; Marsh, E.E., 3rd. Classification of subtype of acute ischemic stroke. Definitions for use in a multicenter clinical trial. TOAST. Trial of Org 10172 in Acute Stroke Treatment. Stroke 1993, 24, 35–41. [Google Scholar] [CrossRef]
- Mehta, R.; Chinthapalli, K. Glasgow coma scale explained. BMJ (Clin. Res. Ed.) 2019, 365, l1296. [Google Scholar] [CrossRef]
- Meng, Q.; Ma, M.; Zhang, W.; Bi, Y.; Cheng, P.; Yu, X.; Fu, Y.; Chao, Y.; Ji, T.; Li, J.; et al. The gut microbiota during the progression of atherosclerosis in the perimenopausal period shows specific compositional changes and significant correlations with circulating lipid metabolites. Gut Microbes. 2021, 13, 1880220. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Gu, M.; Li, Z.; Chen, X.; Zhou, J.J.F.i.n. Gut Microbiota Dysbiosis in Acute Ischemic Stroke Associated With 3-Month Unfavorable Outcome. Front. Neurol. 2021, 12, 799222. [Google Scholar] [CrossRef]
- Li, X.J.; You, X.Y.; Wang, C.Y.; Li, X.L.; Sheng, Y.Y.; Zhuang, P.W.; Zhang, Y.J. Bidirectional Brain-gut-microbiota Axis in increased intestinal permeability induced by central nervous system injury. CNS Neurosci. Ther. 2020, 26, 783–790. [Google Scholar] [CrossRef]
- Wassenaar, T.M.; Juncos, V.A.; Zimmermann, K. Interactions between the Gut Microbiome, Lung Conditions, and Coronary Heart Disease and How Probiotics Affect These. Int. J. Mol. Sci. 2021, 22, 9700. [Google Scholar] [CrossRef]
- Chen, L.W.; Chen, P.H.; Hsu, C.M. Commensal microflora contribute to host defense against Escherichia coli pneumonia through Toll-like receptors. Shock 2011, 36, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Ichinohe, T.; Pang, I.K.; Kumamoto, Y.; Peaper, D.R.; Ho, J.H.; Murray, T.S.; Iwasaki, A. Microbiota regulates immune defense against respiratory tract influenza A virus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 5354–5359. [Google Scholar] [CrossRef]
- Brown, R.; Larkinson, M.; Clarke, T. Immunological design of commensal communities to treat intestinal infection and inflammation. PLOS Pathog. 2021, 17, e1009191. [Google Scholar] [CrossRef]
- Chioma, O.S.; Hesse, L.E.; Chapman, A.; Drake, W.P. Role of the Microbiome in Interstitial Lung Diseases. Front Med. 2021, 8, 595522. [Google Scholar] [CrossRef]
- Nogal, A.; Louca, P.; Zhang, X.; Wells, P.M.; Steves, C.J.; Spector, T.D.; Falchi, M.; Valdes, A.M.; Menni, C. Circulating Levels of the Short-Chain Fatty Acid Acetate Mediate the Effect of the Gut Microbiome on Visceral Fat. Front. Microbiol. 2021, 12, 711359. [Google Scholar] [CrossRef]
- Robert, C.; Chassard, C.; Lawson, P.A.; Bernalier-Donadille, A. Bacteroides cellulosilyticus sp. nov., a cellulolytic bacterium from the human gut microbial community. Int. J. Syst. Evol. Microbiol. 2007, 57, 1516–1520. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.C.; Shen, M.H.; Liu, C.Y.; Pu, C.M.; Hu, J.M.; Huang, C.J. A gut butyrate-producing bacterium Butyricicoccus pullicaecorum regulates short-chain fatty acid transporter and receptor to reduce the progression of 1,2-dimethylhydrazine-associated colorectal cancer. Oncol. Lett. 2020, 20, 327. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhao, G.; Ji, H.; Peng, B.; Huang, Z.; Jin, W.; Chen, X.; Guan, H.; Tang, G.; Zhang, H.; et al. Changes in the gut microbiota during and after commercial helium-oxygen saturation diving in China. Occup. Environ. Med. 2019, 76, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Pattaroni, C.; Lopez-Mejia, I.C.; Riva, E.; Pernot, J.; Ubags, N.; Fajas, L.; Nicod, L.P.; Marsland, B.J. Dietary Fiber Confers Protection against Flu by Shaping Ly6c(-) Patrolling Monocyte Hematopoiesis and CD8(+) T Cell Metabolism. Immunity 2018, 48, 992–1005.e1008. [Google Scholar] [CrossRef] [PubMed]
- Antunes, K.H.; Fachi, J.L.; de Paula, R.; da Silva, E.F.; Pral, L.P.; Dos Santos, A.; Dias, G.B.M.; Vargas, J.E.; Puga, R.; Mayer, F.Q.; et al. Microbiota-derived acetate protects against respiratory syncytial virus infection through a GPR43-type 1 interferon response. Nat. Commun. 2019, 10, 3273. [Google Scholar] [CrossRef]
- Haak, B.W.; Littmann, E.R.; Chaubard, J.L.; Pickard, A.J.; Fontana, E.; Adhi, F.; Gyaltshen, Y.; Ling, L.; Morjaria, S.M.; Peled, J.U.; et al. Impact of gut colonization with butyrate-producing microbiota on respiratory viral infection following allo-HCT. Blood 2018, 131, 2978–2986. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; d’Aigle, J.; Atadja, L.; Quaicoe, V.; Honarpisheh, P.; Ganesh, B.P.; Hassan, A.; Graf, J.; Petrosino, J.; Putluri, N.; et al. Gut Microbiota-Derived Short-Chain Fatty Acids Promote Poststroke Recovery in Aged Mice. Circ. Res. 2020, 127, 453–465. [Google Scholar] [CrossRef]
- Shimizu, K.; Yamada, T.; Ogura, H.; Mohri, T.; Kiguchi, T.; Fujimi, S.; Asahara, T.; Yamada, T.; Ojima, M.; Ikeda, M.; et al. Synbiotics modulate gut microbiota and reduce enteritis and ventilator-associated pneumonia in patients with sepsis: A randomized controlled trial. Crit. Care 2018, 22, 239. [Google Scholar] [CrossRef]
- Benakis, C.; Brea, D.; Caballero, S.; Faraco, G.; Moore, J.; Murphy, M.; Sita, G.; Racchumi, G.; Ling, L.; Pamer, E.G.; et al. Commensal microbiota affects ischemic stroke outcome by regulating intestinal gammadelta T cells. Nat. Med. 2016, 22, 516–523. [Google Scholar] [CrossRef]
- Ott, L. Adhesion properties of toxigenic corynebacteria. AIMS Microbiol. 2018, 4, 85–103. [Google Scholar] [CrossRef]
- Chia, J.H.; Wu, T.S.; Wu, T.L.; Chen, C.L.; Chuang, C.H.; Su, L.H.; Chang, H.J.; Lu, C.C.; Kuo, A.J.; Lai, H.C.; et al. Clostridium innocuum is a vancomycin-resistant pathogen that may cause antibiotic-associated diarrhoea. Clin. Microbiol. Infect. 2018, 24, 1195–1199. [Google Scholar] [CrossRef] [PubMed]
- Highet, A.R.; Berry, A.M.; Bettelheim, K.A.; Goldwater, P.N.J.I.J.o.M.M. Gut microbiome in sudden infant death syndrome (SIDS) differs from that in healthy comparison babies and offers an explanation for the risk factor of prone position. Int. J. Med Microbiol. 2014, 304, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Kuo, Y.C.; Chen, M.C.; Zhang, Y.D.; Chen, C.L.; Le, P.H.; Chiu, C.H. Case-Control Study of Clostridium innocuum Infection, Taiwan. Emerg. Infect. Dis. 2022, 28, 599–607. [Google Scholar] [CrossRef]
- Zanza, C.; Romenskaya, T.; Thangathurai, D.; Ojetti, V.; Saviano, A.; Abenavoli, L.; Robba, C.; Cammarota, G.; Franceschi, F.; Piccioni, A.; et al. Microbiome in Critical Care: An Unconventional and Unknown Ally. Curr. Med. Chem. 2022, 29, 3179–3188. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, K.; Li, Z.; Chai, X.; Fu, X.; Kholodkevich, S.; Kuznetsova, T.; Chen, C.; Ren, N. Effescts of acute diclofenac exposure on intestinal histology, antioxidant defense, and microbiota in freshwater crayfish (Procambarus clarkii). Chemosphere 2021, 263, 128130. [Google Scholar] [CrossRef]
- García-Solache, M.; Rice, L.B. The Enterococcus: A Model of Adaptability to Its Environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Chen, Y.; Tang, G.; Li, Z.; Yang, X.; Shang, X.; Huang, T.; Huang, G.; Wang, L.; Han, Y.; et al. Gut microbiota composition reflects disease progression, severity and outcome, and dysfunctional immune responses in patients with hypertensive intracerebral hemorrhage. Front. Immunol. 2022, 13, 869846. [Google Scholar] [CrossRef]
- Liu, D.D.; Chu, S.F.; Chen, C.; Yang, P.F.; Chen, N.H.; He, X. Research progress in stroke-induced immunodepression syndrome (SIDS) and stroke-associated pneumonia (SAP). Neurochem. Int. 2018, 114, 42–54. [Google Scholar] [CrossRef]
- Bourassa, M.W.; Alim, I.; Bultman, S.J.; Ratan, R.R.J.N.L. Butyrate, neuroepigenetics and the gut microbiome: Can a high fiber diet improve brain health? Neurosci. Lett. 2016, 625, 56–63. [Google Scholar] [CrossRef]
- Chen, T.; Chen, X.; Zhang, S.; Zhu, J.; Tang, B.; Wang, A.; Dong, L.; Zhang, Z.; Yu, C.; Sun, Y.; et al. The Genome Sequence Archive Family: Toward Explosive Data Growth and Diverse Data Types. Genom. Proteom. Bioinform. 2021, 19, 578–583. [Google Scholar] [CrossRef]
- CNCB-NGDC. Database Resources of the National Genomics Data Center, China National Center for Bioinformation in 2021. Nucleic Acids Res. 2021, 49, D18–D28. [Google Scholar] [CrossRef] [PubMed]