


1. Introduction
1.1. Ovarian Cancer
Ovarian cancer (OC) is a lethal gynecological malignancy. According to the GLOBOCAN 2020 statistics, there were 314,000 new cases and 207,000 deaths in the world, projecting a 100% increase in OC deaths by 2040 [1]. Epithelial OC (EOC) is the most common subtype of OCs (representing over 95% of OC) and is characterized by non-specific symptoms and late diagnosis, which results in poor prognosis. High-grade serous epithelial OC (HGS-EOC) is very aggressive and represents 75% of OC [2,3].
From a molecular point of view, inflammation and angiogenesis (the formation of new blood vessels from pre-existing ones) are two important and connected processes involved in the progression, chemoresistance, and dissemination of OC [4,5]. Chronic inflammation is strictly related to cancer risk, producing an increase in cell proliferation and reduced DNA repair [6]. Both extrinsic and intrinsic inflammation (produced by the immune system and by the own cancer cells, respectively) contribute to multiple hallmark capabilities by supplying bioactive molecules to the tumor microenvironment, including growth factors that sustain proliferative signaling, survival factors that limit cell death, proangiogenic factors, extracellular-matrix-modifying enzymes that facilitate angiogenesis, invasion, and metastasis, and inductive signals that lead to the activation of epithelial-mesenchymal transition (EMT) [7]. In EOC, increased expression of cyclooxygenase (COX)-1 and COX-2, and the increase in inflammatory mediators, such as prostaglandin E2 (PGE2), have been associated with poor prognosis [8,9,10]. The inflammatory signaling promotes the survival of cancer cells and causes genomic instability, allowing mutant cells to escape cell cycle arrest and apoptosis [11].
Another key characteristic of EOC is its high angiogenesis potential, since EOC cells produce several angiogenic factors, such as the vascular endothelial growth factor (VEGF) family, angiopoietins, neurotrophins, fibroblast growth factors (FGF), and platelet-derived growth factor (PDGF), among others [12,13]. This knowledge has served to develop several anti-angiogenic therapies that are being tested in patients with OC [12], including Bevacizumab, a monoclonal antibody against VEGF-A that was approved by the FDA in 2018 for use in patients with advanced-stage OC.
1.2. Current and New Approaches for OC
The current treatment for EOC is debulking surgery. Primary cytoreductive surgery followed by adjuvant chemotherapy remains the standard treatment for EOC in advanced stages [14]. Despite optimal surgery and chemotherapy, ∼70–80% of patients with EOC will develop disease relapse [14,15], indicating the need to improve the existing therapies.
New therapeutic alternatives have been studied for OC; some of them are natural compounds, such as flavonoids and polyphenols, which have shown antitumoral effects in several kinds of cancer, including OC [16]. Other natural compounds, such as withanolides (bioactive molecules isolated from Whitania somnifera or Indian ginseng), have shown enhanced therapeutic activity as anticancer drugs, suppressing growth and metastasis in OC models [17,18]. In this line, the design of small peptides that inhibit mutant oncogenic proteins, such as K-Ras, one of the major gene mutations correlated with OC occurrence [19], has promising results [20]. On the other hand, the use of repurposing drugs (or the investigation of approved drugs for new therapeutic purposes, such as cancer treatment) is being considered as a therapeutic approach in OC [14]. Thus, drugs such as metformin have been studied in in vitro and in vivo models and are currently being tested in several clinical trials as a complementary therapy for OC.
1.3. Metformin in OC
Evidence from retrospective studies has shown that the use of some drugs, such as metformin, could decrease OC incidence and mortality in diabetic patients [21]. Metformin is a widely used drug for the treatment of metabolic disorders, such as type 2 diabetes mellitus, metabolic syndrome, gestational diabetes, and polycystic ovarian syndrome [22,23]. Metformin produces normoglycemia by decreasing hepatic glucose production, the intestinal absorption of glucose, and improving insulin sensitivity, thus promoting glucose uptake and utilization by peripheral tissues [22]. Metformin enters the cell through organic cation transporters (OCTs) and multidrug and toxin extrusion transporters [24,25] and is accumulated in tissues that express OCT transporters, such as the ovary [26,27,28], which are, therefore, adequate targets for metformin action. In the context of cancer, the direct antitumoral effects of metformin are pleiotropic and involve several molecular targets at different levels in the tumoral cell, including epigenetic regulation (changes in cell function without alterations in the DNA sequence) [29].
In vitro experiments showed that metformin decreased EOC cell proliferation by inducing cell cycle arrest and altering glucose and lipid metabolism [30,31]. In vivo experiments showed that metformin treatment decreased OC growth and the presence of cell proliferation markers (such as Ki-67 and cyclin D1), as well as metastasis [32,33]. Most of these antitumoral effects exerted by metformin depended on the activation of the enzyme 5′ adenosine monophosphate-activated protein kinase (AMPK), which induces p53 phosphorylation (S15), which is required for cell cycle arrest [34]. In addition, high doses of metformin inhibited the mitochondrial respiratory chain complex I, activating AMPK, increasing reactive oxygen species (ROS), and producing a glycolytic switch in OC cells [35].
In addition to its anti-proliferative effects, metformin treatment inhibits both angiogenesis and metastatic spread in OC models [32,33]. In vitro and in vivo studies showed that mechanistic inhibition of angiogenesis by metformin involves the inhibition of the expression of angiogenic factors, such as VEGF, as well as a decrease in endothelial cell migration [36] and the polarization of tumor-associated macrophages (TAMs) [37].
On the other hand, metformin has been described to prevent the EMT phenotype in vitro [38], which is associated with cell cycle arrest and the accumulation of OC cells in the S-phase of the cell cycle [39]. In agreement with the last point, several in vitro and in vivo studies have shown that platinum or taxane therapy allows OC cells to acquire a mesenchymal phenotype [40,41,42]. EMT proteins such as Snail, Slug, Twist-1, and Zeb-1 contribute not only to OC dissemination but also increase cisplatin and carboplatin resistance in OC cells [41,43,44,45]. In addition to these antecedents, evidence suggests that metformin targets cancer stem cells: a phase II clinical trial showed that metformin decreased the population of aldehyde dehydrogenase (ALDH)(+) and CD33(+) cells in ovarian tumors [46]. All of this evidence could explain the inhibitory effects of metformin on the metastatic behavior of OC cells.
Metformin has pleiotropic effects and targets OC cells at multiple levels. Since non-coding RNAs (ncRNAs) can regulate several proteins and produce multiple changes at cellular levels, it is plausible that the antitumoral effects of metformin in OC could be explained by ncRNA regulation. In fact, recent evidence has shown that metformin treatment could regulate the expression of ncRNAs such as microRNAs (miRNAs) and long non-coding RNAs (lncRNAs) to exert antitumoral effects.
Few studies have shown that the antitumor effects of metformin can be attributed directly, or at least in part, to the modulation of ncRNAs in OC. In this review, we discuss the existing evidence regarding how metformin could modulate the levels of some important ncRNAs in OC. Since the studies are limited, we decided to discuss a possible connection between the changes in some ncRNAs in OC progression and their modulation by metformin, with a focus on miRNAs and lncRNAs.
2. Methodological Approach: Search Strategy and Articles Inclusion Criteria
We searched the PubMed database for articles that described the effects of metformin on cancer cells and whose antitumoral mechanisms are related to the modulation of ncRNAs. Articles were published between 2006 and 2023. The following search terms were used: (“metformin”) AND (“cancer” OR “carcinoma”) AND ((“noncoding RNAA”; OR “ncRNA”) OR (“microRNA” OR “miR”) OR (“long noncoding RNA”; OR “lncRNA”) OR (“circRNA” OR “circular RNA”)). A total of 255 potential studies were found through database searching, and an initial screening was performed, including only articles written in English.
The PubMed database was also used to search for articles describing relevant oncogenic ncRNAs in OC that are regulated by metformin, as indicated in the former articles found. The following search terms were used: “miR-21” AND (“ovarian cancer”; OR “ovarian carcinoma”); “miR-27a” AND (“ovarian cancer”; OR “ovarian carcinoma”); “H19” (long non-coding RNA H19) AND (“ovarian cancer”; OR “ovarian carcinoma”); “SNHG7” (mall Nucleolar RNA Host Gene 7) AND (“ovarian cancer”; OR “ovarian carcinoma”). The number of articles found related to relevant oncogenic ncRNAs in OC was 88 (miR-21), 22 (miR-27a), 39 (H19), and 4 (SNHG7).
To discuss and propose the new ncRNAs that metformin could potentially regulate in OC, we considered those articles from the initial screening that explored modulated ncRNAs by metformin in cancer cells, including OC cells, according to the following criteria: (1) the articles describe metformin-regulated ncRNAs that are considered oncogenic in OC, and (2) ncRNAs that are downregulated by metformin. We further considered these oncogenic ncRNAs according to their relevance in OC development and progression, and the number of articles that resulted from the PubMed database search.
3. Non-Coding RNAs
Non-coding RNAs (ncRNAs) play a key role in the pathophysiology of many diseases, including cancer. There are different kinds of ncRNAs: transcripts of less than 200 nt are small non-coding RNAs, while transcripts bigger than 200 nt are lncRNAs, of which miRNAs are the main class of ncRNAs [47]. The regulation of the expression of ncRNAs by metformin may involve epigenetic modifications (for instance, modification of DNA methylation) as well as transcription regulation and modulation of maturation.
Figure 1 shows the diagram of miRNA synthesis, which begins with the transcription of a primary miRNA (pri-miRNA). Then, an imperfect hairpin structure is cleaved, producing a smaller hairpin called precursor miRNA (Pre-miRNA). It is exported from the nucleus to the cytoplasm through exportin 5, a double-stranded RNA-binding protein. The Pre-miRNA is processed by the RNase III Dicer, a double-stranded RNA nuclease, producing a small miRNA duplex. In the next step, the miRNA duplex (miR duplex) binds Argonaute (AGO) proteins, a family of gene-silencing proteins guided by small RNAs, such as miRNA, that get into specific binding pockets and guide AGO proteins to target messenger RNAs (mRNA) [48]. After AGO ejects the passenger strand (a strand that will not be used to form the silencing complex), the guide strand bound to AGO, along with other proteins, will form the RNA-induced silencing complex (RISC), which will produce the degradation of the mRNA of complementary sequence to the miRNA or the inhibition of its translation [49].
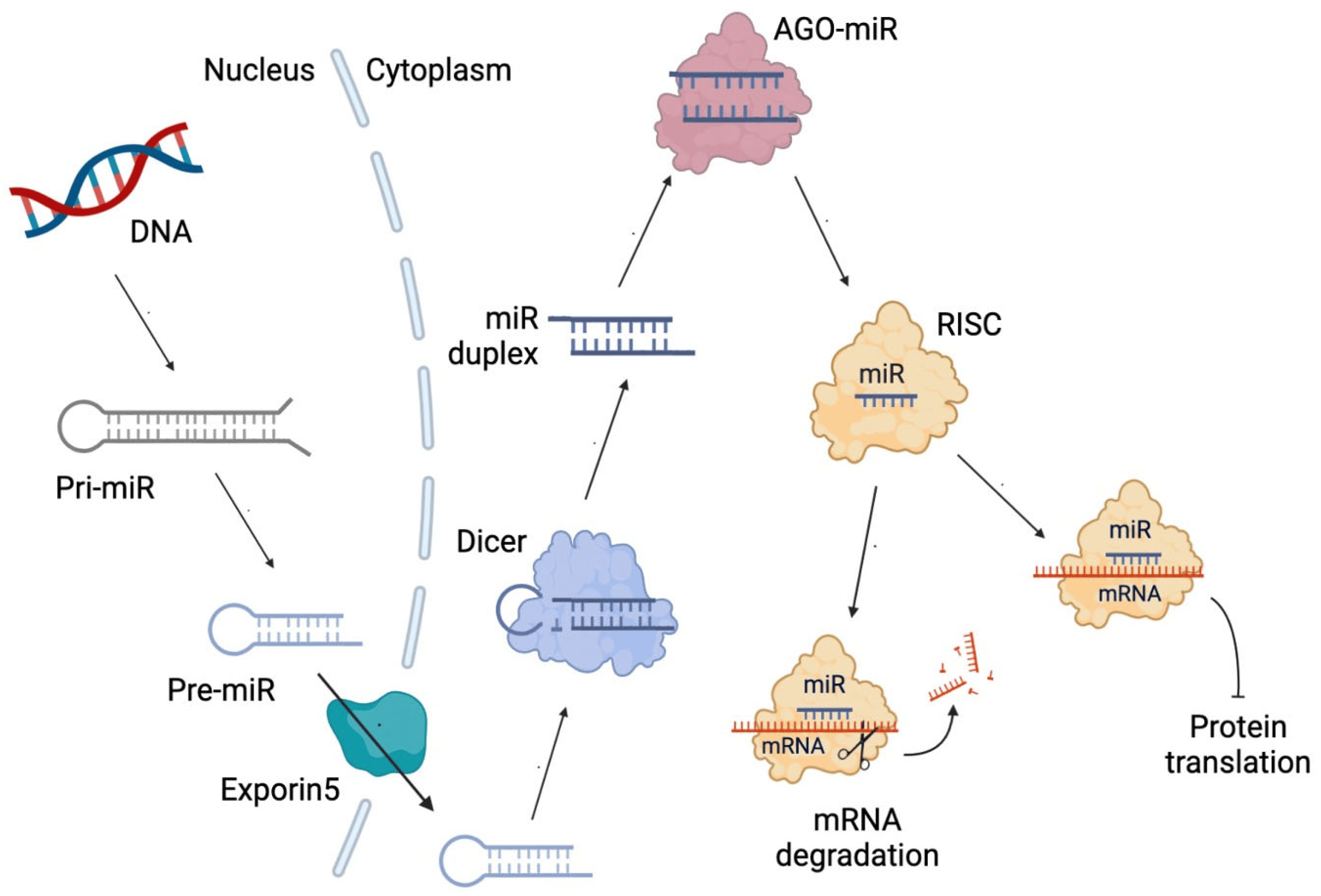
Figure 1. Diagram of microRNA (miRNA) biosynthesis. miR duplex: miRNA duplex comprising two strands. pri-miRNA: primary miRNA. Pre-miRNA: precursor miR. mRNA: messenger RNA. Dicer: RNase III double-stranded RNA nuclease. AGO-miR: Argonaute bound to miRNA. RISC: RNA-induced silencing complex.
4. Mechanisms of ncRNA Biosynthesis Regulated by Metformin
The pleiotropic anticancer effects of metformin have been studied recently through multiple pathways, including its ability to regulate the expression of ncRNAs [50]. Metformin modulates the expression of several ncRNAs in different cancer cells, including OC cells, affecting their tumor formation and progression capacity [51]. However, most studies have focused on describing changes in ncRNA levels upon metformin treatment, and very few have explored the regulatory mechanism by which metformin alters the expression of these ncRNAs. To help build a more in-depth evidence framework, in this section, we discuss the previously described mechanisms through which metformin manages to alter the expression of ncRNAs, with a focus on OC and miR-145.
4.1. Role of Transcriptional Factors in the Metformin-Mediated Modulation of miRNA Expression
4.2. Role of Metformin on Epigenetic Modification of ncRNAs
4.3. Role of Metformin in miRNA Maturation
Several studies have suggested that post-transcriptional maturation, rather than transcription, is often altered in cancer [88]. Accordingly, pri-miRNAs accumulate and deplete mature miRNAs in human cancer [89], indicating that the machinery for processing and maturation of miRNAs is dysregulated in cancer. Notably, metformin has been shown to induce the expression of key processing enzymes of miRNA biogenesis. Metformin treatment upregulates Drosha and Dicer expression in cholangiocarcinoma and breast cancer cells, respectively, to modulate miRNA expression [66,90]. On the other hand, decreased Dicer and Drosha levels represent an oncogenic event in EOC cells and are associated with poor patient outcomes [91,92,93]. Interestingly, p70S6 kinase (p70S6K), a downstream effector of phosphoinositide 3-kinases (PI3K)/protein kinase B (Akt)/mechanistic target of rapamycin (mTOR) signaling, was shown to affect the miRNA biogenesis machinery in EOC cells [94]. P70S6K phosphorylates tristetraprolin (TTP), preventing its interaction with Dicer, which specifically promotes the maturation of miR-145. Metformin may also regulate miRNA expression by influencing the activity of miRNA processing enzymes in OC. As reported, metformin impairs PI3K/Akt/mTOR signaling and thus decreases the active phosphorylated form of P70S6K in EOC cells [58,95,96]. Since metformin can increase miR-145 levels in EOC cells [80], these findings suggest that metformin may prevent p70S6K activation to favor miR-145 expression through Dicer and TTP interaction in EOC cells.
5. ncRNA-Related Therapeutic Effects of Metformin in OC
Very few studies have reported that metformin’s anticancer effects may involve the regulation of ncRNAs in OC. Together with the studies that our group has carried out regarding the regulation of tumor suppressors miR-145 and miR-23b by metformin [80], only two additional studies reported an influence of metformin on ncRNAs in OC [71,75]. These studies showed that treatment of OC cells with metformin reduced the expression of the lncRNAs H19 and SNHG7, which was associated with anticancer effects. Because there are few studies related to this topic in OC, in this section we describe some important ncRNAs in the pathogenesis and progression of OC and discuss their potential as relevant metformin targets.
5.1. miR-23b and miR-145 in OC
miR-23b and miR-145 are two oncosupressor miRNAs that are downregulated in EOC cell lines and EOC tissues, as we have reported [97]. To date, our group has shown that metformin increases the expression levels of tumor suppressors miR-145 and miR-23b in EOC cells and prevents the nerve growth factor (NGF)-induced decrease in these miRNAs [80]. These changes coincided temporarily with a decrease in the expression and transcriptional activity of c-MYC upon metformin treatment. Other authors have shown that c-MYC can reduce the expression of both miR-145 and miR-23b in cancer [98,99]. Furthermore, a study performed with a small sample of EOC patients showed that metformin intake reduced the presence of oncoproteins related to cell proliferation, such as c-MYC and survivin [80]. Consistent with these findings, we have further shown that NGF reduces the transcriptional activation of the miR-145 promoter while inducing c-MYC protein levels in EOC cells [68]. Therefore, NGF could repress miR-145 and miR-23b, and these effects could be mediated by c-MYC activation, while in the presence of metformin, this NGF ability is impaired. These observations suggest that metformin potentially modulates transcription factors, such as c-MYC, to increase tumor suppressor miRNA levels in OC cells.
5.2. miR-21 in OC and Other Cancers
miR-21 is considered a pro-tumoral miRNA in EOC since its overexpression increases cell proliferation, invasion, and migration abilities of EOC, and decreases apoptotic cell death in EOC tumors [100,101,102,103]. Additionally, miR-21-5p is involved in paclitaxel resistance in EOC cells because it can sensitize OC cells to paclitaxel, reducing cell proliferation, migration, invasion, and EMT [104].
Although there is vast evidence of the pro-tumoral role of miR-21 in EOC, and metformin has been shown to downregulate this miRNA in several kinds of cancer, the direct effect of metformin on miR-21 in EOC has not been studied yet. Nevertheless, a connection must exist between metformin and miR-21 since metformin downregulates this miRNA in a diversity of human cancer models. For instance, a reduction of miR-21 levels in breast cancer cells following metformin treatment has been assessed both in vitro and in vivo [65,105]. Also, in chemo-resistant colon cancer cells (highly enriched in cancer stem cells), metformin causes a marked reduction of miR-21 expression, and this effect is further accomplished in combination with oxaliplatin treatment [106]. Metformin has been shown to act synergistically with oxaliplatin to induce cell death, inhibit colonosphere formation and cell migration, and inhibit tumor growth of colon cancer cells. In the case of renal cancer cells, metformin treatment can decrease miR-21 expression and increase PTEN levels, impairing PI3K/Akt signaling and cell proliferation [107]. A similar effect has been shown in hypopharyngeal cancer cells, in which metformin treatment inhibited miR-21 expression, causing a decrease in cell proliferation [54].
Additionally, metformin has shown anti-angiogenic effects in EOC cells; it improves OC sensitivity to paclitaxel and platinum-based drugs and decreases the metastatic potential of OC cells [31,80,108]. Since the upregulation of miR-21 has been described in these processes [109,110], it is possible that these effects could be mediated by a metformin-dependent decrease in miR-21.
5.3. miR-27 in OC and Other Cancers
Another important miRNA is miR-27a, which represents an important oncomiR involved in OC development, progression, and chemoresistance. It stimulates cell proliferation, cell cycle progression, migration, invasion, and EMT of EOC cells by directly targeting a tumor suppressor transcription factor, forkhead box O1 (FOXO1). Metformin has been shown to downregulate miR-27a in cancer cells of several origins. For example, in breast cancer cells, metformin inhibits growth and promotes apoptosis by decreasing miR-27a levels [111]. Additionally, the expression of miR-27a is upregulated in taxol-resistant EOC cells, which in turn could increase multidrug-resistant 1 (MDR1) expression. Since miR-27a could directly target AMPK [111], metformin could activate AMPK by downregulating miR-27a in cancer cells. On the other hand, miR-27a levels decreased upon metformin treatment in pancreatic cancer cells, upregulating transcription factor repressors (zinc finger and BTB domain-containing protein 10 (ZBTB10)) [112] of the specificity protein (Sp). This leads to downregulation of Sp1/3/4 transcription factor and several pro-angiogenic Sp-regulated genes, including bcl-2, survivin, cyclin D1, VEGF, VEGF receptor, and fatty acid synthase, which may account for the antitumoral effects of metformin in pancreatic tumor growth. The effect of metformin on miR-27a has not been assessed yet in OC. Since miR-27a is also an important oncomiR that has overlapping target genes with miR-21, its downregulation could explain in part the anti-angiogenic, anti-metastatic, and chemo-sensitizing effects of metformin in OC.
In summary, solid scientific evidence has shown that metformin can regulate miRNA abundance, such as miR-145 and miR-23b, in EOC. On the other hand, metformin regulates miR27a and miR21 in different kinds of cancer, suggesting that metformin could be implicated in the regulation of these miRNAs in OC. However, metformin not only modulates the expression and abundance of miRNAs but also lncRNAs and Figure 2). In fact, a wide variety of these ncRNAs can be regulated by metformin. Next, we will review the evidence of the effects of metformin on two very important lncRNAs in EOC: H19 and SNHG7.
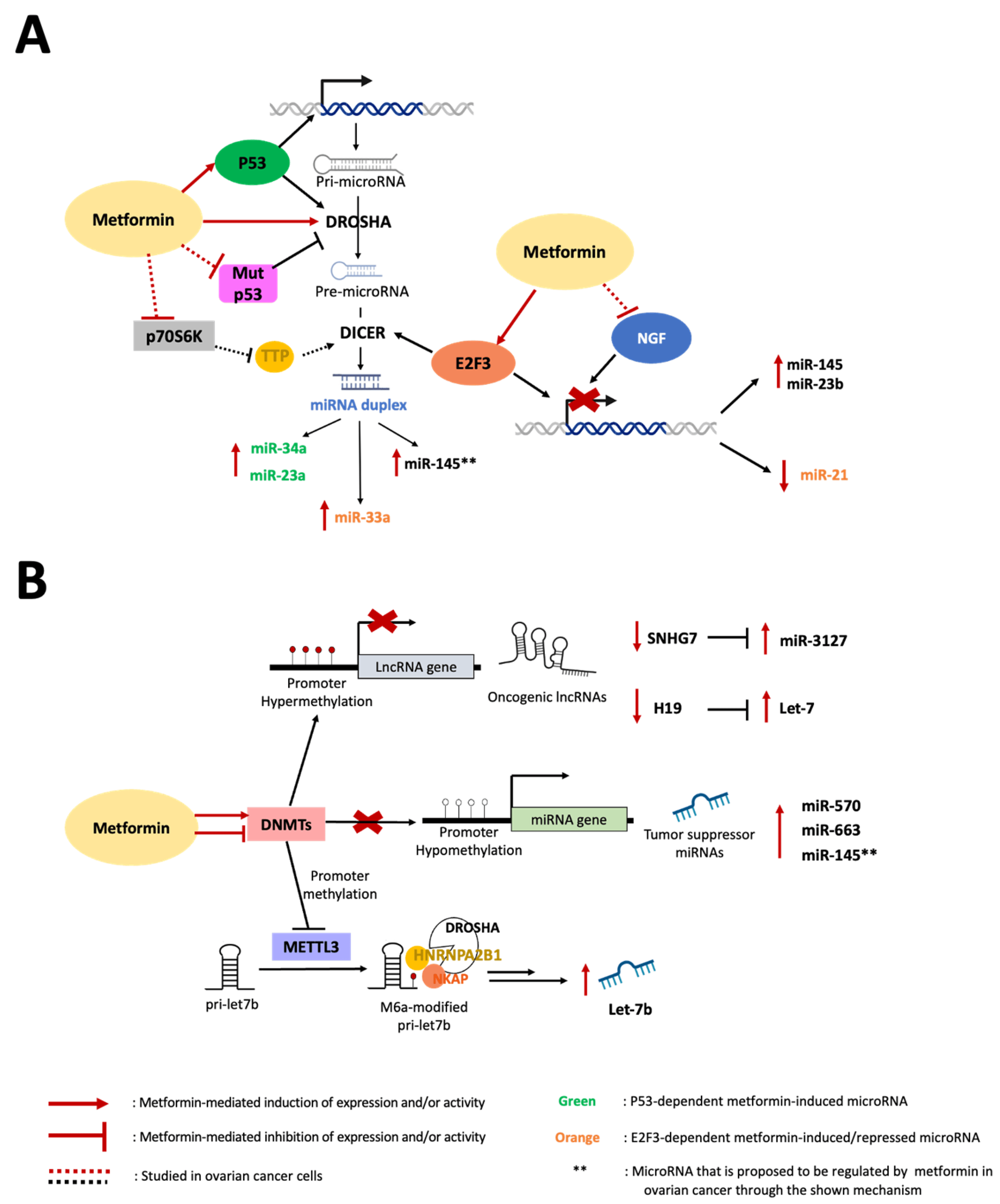
Figure 2. Mechanisms of ncRNA biosynthesis by metformin. (A) Metformin induces the expression and activity of wild-type p53 to upregulate miR-34a and miR-23a in carcinoma cells and represses miR-21 expression by promoting the activity of E2F3 on the miR-21 promoter while inducing miR-33a by E2F3-mediated upregulation of DICER. In EOC cells, metformin prevents the NGF-induced decrease in miR-145 and miR-23b and increases the expression levels of these miRNAs. We propose that miR-145 is upregulated by metformin in EOC cells through the induction of p53 (which regulates transcription and maturation), inhibition of mutant p53 expression (which disfavors Drosha activity on pri-miR-145), and Akt/mTOR/p70S6K signaling impairment (which hinders pre-miR-145 processing by Dicer). (B) In EOC cells, metformin treatment induces hypermethylation in the H19 and SNHG7 promoters, increasing the bioavailability of let-7 and miR-3127 by releasing them from lncRNA sequestration. Metformin-mediated SNHG7 promoter hypermethylation has been associated with DNMT induction in hypopharyngeal cancer cells. In contrast, metformin treatment produces miR-663 and miR-570 promoter hypomethylation (upregulating their expression) in pancreatic and osteosarcoma cancer cells, respectively. We propose that miR-145 is upregulated by metformin through hypomethylation of its promoter in EOC cells. Metformin inhibits DNMTs binding to the METTL3 promoter, lowering its methylation status, and increases METTL3 expression, allowing m6A modification of pri-let-7b, which binds m6A reader factors (HNRNPA2B1 and NKAP). These produce Dicer recognition and processing of pri-let-7b into mature let-7b.
5.4. lncRNA H19 in EOC and Its Regulation by Metformin
H19 is a lncRNA widely considered an oncogene in various types of cancer, including EOC, and it is critically involved in tumor development, malignant progression, and chemoresistance [125]. H19 is highly expressed in human ovarian tumor tissues and has been associated with cancer progression and poor patient prognosis [126,127]. Moreover, high levels of H19 can also be detected in ascite fluids from OC patients [128], suggesting a role in metastasis development. Knockdown of H19 has been shown to decrease cell proliferation, migration, and invasion in OC cells [129,130,131]. In addition, H19 acts as a molecular sponge of miRNAs [127,132], and controls OC metabolism (favoring the Warburg effect) [133]. Moreover, H19 also participates in the development of chemoresistance in EOC. H19 can enhance the chemoresistance of OC cells to carboplatin by antagonizing miR-29, which increases multidrug resistance proteins, such as MDR1 and MRP1 [134]. LncRNA H19 contributes to enhancing the growth and cell cycle of cancers and inducing EMT and, therefore, promotes metastasis [135]. Importantly, the knockdown of H19 in cisplatin-resistant EOC cells improves cisplatin sensitivity in vitro and in vivo through glutathione metabolism impairment [136]. This report indicated that H19 is involved in tumor development, malignant progression, and resistance to chemotherapy in EOC, and, therefore, it profiles as a relevant therapeutic target.
In terms of how metformin regulates H19 in EOC cells, Yan et al. demonstrated that metformin decreases H19 expression by altering its promoter methylation, inhibiting EOC cell migration and invasion [71]. Accordingly, it was previously shown that histone H1.3 directly represses the expression of the H19 gene in EOC cells through a similar epigenetic mechanism involving DNA methylation [137]. Downregulation of H19 relieves let-7 inhibition from H19 sponging, leading to decreased metastasis-promoting genes such as c-MYC, the high-mobility group AT-hook 2 (HMGA2), and U3 small nucleolar ribonucleoprotein (IMP3) [71]. Although evidence suggests that the use of metformin might be useful in targeting H19 in OC therapy, these findings are limited to cell lines, and further studies are needed to test this metformin effect in vivo. Nevertheless, according to the above, studies in other cancer types have supported the idea that metformin could be clinically useful in targeting H19 and producing antitumor effects. For instance, endometrial cancer patients receiving antidiabetic doses of metformin have reduced H19 expression levels in the endometrial tumor tissue [82,120]. In preclinical studies, metformin treatment has been shown to inhibit cell migration and invasion by downregulating H19 expression in gastric cancer cells [121]. Furthermore, in a mouse model injected intravenously with gastric cancer cells, metformin suppressed metastasis in a similar way to that of mice that were injected with H19-knockdown cells. Similarly, a recent study conducted in breast cancer cells showed that metformin may induce ferroptosis by downregulating H19 [116]. All this evidence shows that metformin decreases H19 in different cancers, and the pivotal role of H19 in EOC supports further metformin research as a relevant H19-targeting therapeutic agent in OC.
5.5. Metformin Regulation of lncRNA SNHG7 in OC
The lncRNA SNHG7, along with the lncRNA H19, is one of the well-studied oncogenes involved in the development of multiple cancers [138]. However, its study in EOC is rather scarce. In ovarian tumor tissues, SNHG7 expression was shown to be elevated [139], suggesting its role as an oncogene. Consistently, the knockdown of SNHG7 in OC cells decreased their growth, migration, and invasion abilities and inhibited tumorigenesis in vivo [139]. Additionally, silencing SNHG7 expression enhanced the sensitivity of resistant OC cells to paclitaxel and reduced their migrative and invasive potential [140].
A recent study reported for the first time that metformin enhances paclitaxel sensitivity and inhibits cell viability, migration, and invasion by decreasing SNHG7 expression in paclitaxel-resistant OC cells [75]. Simultaneously, metformin was shown to promote miR-3127 expression, which proved to be a direct target of SNHG7, which acts as a molecular sponge. Importantly, the regulatory effect of metformin on the SNHG7/miR-3127 ratio was confirmed in a xenograft model of OC, in which metformin prevented the promotion of tumor growth by SNHG7 overexpression [75]. In addition, metformin can also downregulate SNHG7 expression in hypopharyngeal cancer cells to inhibit cell proliferation and improve paclitaxel sensitivity [141]. There is still very little evidence of the SNHG7 oncogenic role and its regulation by metformin as an anticancer mechanism in OC. However, we hope that the existing results will encourage further research as metformin increasingly continues to profile as an attractive therapeutic alternative in OC.
6. Future Directions
The antitumoral effects of metformin, as evidenced in in vitro and retrospective studies, have supported the study of metformin in non-diabetic patients with different kinds of cancer. There are more than 400 clinical trials registered in ClinicalTrials.gov that have tested metformin in patients with and without diabetes with different kinds of cancer. Several of these trials yielded results; however, the results are contradictory. For instance, in patients with breast cancer cells, the use of metformin vs. placebo did not significantly improve invasive disease-free survival in the entire arm. However, ERBB2+ patients had longer invasive disease-free survival [142]. Similarly, a study of patients with advanced pancreatic cancer showed that metformin does not improve outcomes in the group treated with gemcitabine, erlotinib, and metformin, but the overall survival was significantly longer in the 16 patients with higher concentrations of metformin [143]. In this line, a study performed on patients with breast cancer shows two different patterns: metformin responders and non-responders, suggesting that patients with an increase in oxidative phosphorylation gene transcription could be resistant to metformin treatment [144].
However, there are some pilot studies that tested the effects of metformin in patients without diabetes or metabolic abnormalities in a “preoperative window” or in patients with pre-malignant lesions, which show more consistent results. summarizes some of these studies and their main findings.
Taking this into consideration, the different metformin responses observed in several studies could be a consequence of the different metformin concentrations allowed in blood or tissue, the presence of specific mutations, or a metabolic signature. We think that a miRNA profile could be useful to characterize patients who may respond better to metformin treatment. Even though almost every clinical trial that tested metformin in cancer patients did not consider the study of the miRNA profile (enrolled in ClinicalTrials.gov), some of them attempted to measure changes in miRNAs, which are listed in . The information provided by these studies can be very valuable in clarifying the differential response observed in cancer patients treated with metformin.
7. Conclusions
Increasing scientific and clinical evidence supports the antitumor effects of metformin in cancer, including OC. Several of its antitumoral effects can be explained as multiple effects at the post-transcriptional level, which include the modulation of miRNAs and lncRNAs studied in different models of cancer. Some possible mechanisms that could be implicated in the antitumoral effects of metformin in OC are the upregulation of miR-145 and miR-23b and the downregulation of miR-21 and miR-27a. Furthermore, metformin could downregulate some lncRNAs, such as H19 and SNHG7, which in turn could modulate miRNA expression and many protein targets of metformin. However, the evidence for the regulation of ncRNAs by metformin in OC is still scarce and needs to be further investigated.
References
- Cabasag, C.J.; Fagan, P.J.; Ferlay, J.; Vignat, J.; Laversanne, M.; Liu, L.; van der Aa, M.A.; Bray, F.; Soerjomataram, I. Ovarian cancer today and tomorrow: A global assessment by world region and Human Development Index using GLOBOCAN 2020. Int. J. Cancer 2022, 151, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Yancik, R. Ovarian cancer. Age contrasts in incidence, histology, disease stage at diagnosis, and mortality. Cancer 1993, 71, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Arora, T.; Mullangi, S.; Lekkala, M.R. Ovarian Cancer. In StatPearls; StatPearls Publishing LLC: Treasure Island, FL, USA, 2023. [Google Scholar]
- Savant, S.S.; Sriramkumar, S.; O’Hagan, H.M. The Role of Inflammation and Inflammatory Mediators in the Development, Progression, Metastasis, and Chemoresistance of Epithelial Ovarian Cancer. Cancers 2018, 10, 251. [Google Scholar] [CrossRef] [PubMed]
- Gavalas, N.G.; Liontos, M.; Trachana, S.P.; Bagratuni, T.; Arapinis, C.; Liacos, C.; Dimopoulos, M.A.; Bamias, A. Angiogenesis-related pathways in the pathogenesis of ovarian cancer. Int. J. Mol. Sci. 2013, 14, 15885–15909. [Google Scholar] [CrossRef]
- Guo, Y.; Nie, Q.; MacLean, A.L.; Li, Y.; Lei, J.; Li, S. Multiscale Modeling of Inflammation-Induced Tumorigenesis Reveals Competing Oncogenic and Oncoprotective Roles for Inflammation. Cancer Res. 2017, 77, 6429–6441. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Beeghly-Fadiel, A.; Wilson, A.J.; Keene, S.; El Ramahi, M.; Xu, S.; Marnett, L.J.; Fadare, O.; Crispens, M.A.; Khabele, D. Differential cyclooxygenase expression levels and survival associations in type I and type II ovarian tumors. J. Ovarian Res. 2018, 11, 17. [Google Scholar] [CrossRef]
- Daikoku, T.; Wang, D.; Tranguch, S.; Morrow, J.D.; Orsulic, S.; DuBois, R.N.; Dey, S.K. Cyclooxygenase-1 is a potential target for prevention and treatment of ovarian epithelial cancer. Cancer Res. 2005, 65, 3735–3744. [Google Scholar] [CrossRef]
- Ye, Y.; Wang, X.; Jeschke, U.; von Schonfeldt, V. COX-2-PGE(2)-EPs in gynecological cancers. Arch. Gynecol. Obstet. 2020, 301, 1365–1375. [Google Scholar] [CrossRef]
- Pua, K.H.; Chew, C.L.; Lane, D.P.; Tergaonkar, V. Inflammation-associated genomic instability in cancer. Genome Instab. Dis. 2020, 1, 1–9. [Google Scholar] [CrossRef]
- Mei, C.; Gong, W.; Wang, X.; Lv, Y.; Zhang, Y.; Wu, S.; Zhu, C. Anti-angiogenic therapy in ovarian cancer: Current understandings and prospects of precision medicine. Front. Pharmacol. 2023, 14, 1147717. [Google Scholar] [CrossRef]
- Garrido, M.P.; Torres, I.; Vega, M.; Romero, C. Angiogenesis in Gynecological Cancers: Role of Neurotrophins. Front. Oncol. 2019, 9, 913. [Google Scholar] [CrossRef]
- Garrido, M.P.; Fredes, A.N.; Lobos-Gonzalez, L.; Valenzuela-Valderrama, M.; Vera, D.B.; Romero, C. Current Treatments and New Possible Complementary Therapies for Epithelial Ovarian Cancer. Biomedicines 2021, 10, 77. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Ueda, Y.; Naka, T.; Enomoto, T. Therapeutic strategies in epithelial ovarian cancer. J. Exp. Clin. Cancer Res. 2012, 31, 14. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, L.; Nandy, S.K.; Payal, N.; Yadav, S.; Vargas-De-La-Cruz, C.; Anwer, M.K.; Khan, H.; Behl, T.; Bungau, S.G. Molecular Aspects and Therapeutic Implications of Herbal Compounds Targeting Different Types of Cancer. Molecules 2023, 28, 750. [Google Scholar] [CrossRef] [PubMed]
- Sivasankarapillai, V.S.; Madhu Kumar Nair, R.; Rahdar, A.; Bungau, S.; Zaha, D.C.; Aleya, L.; Tit, D.M. Overview of the anticancer activity of withaferin A, an active constituent of the Indian ginseng Withania somnifera. Environ. Sci. Pollut. Res. Int. 2020, 27, 26025–26035. [Google Scholar] [CrossRef] [PubMed]
- Kakar, S.S.; Ratajczak, M.Z.; Powell, K.S.; Moghadamfalahi, M.; Miller, D.M.; Batra, S.K.; Singh, S.K. Withaferin a alone and in combination with cisplatin suppresses growth and metastasis of ovarian cancer by targeting putative cancer stem cells. PLoS ONE 2014, 9, e107596. [Google Scholar] [CrossRef] [PubMed]
- Therachiyil, L.; Anand, A.; Azmi, A.; Bhat, A.; Korashy, H.M.; Uddin, S. Role of RAS signaling in ovarian cancer. F1000Research 2022, 11, 1253. [Google Scholar] [CrossRef]
- Ghufran, M.; Khan, H.A.; Ullah, M.; Ghufran, S.; Ayaz, M.; Siddiq, M.; Hassan, S.S.U.; Bungau, S. In Silico Strategies for Designing of Peptide Inhibitors of Oncogenic K-Ras G12V Mutant: Inhibiting Cancer Growth and Proliferation. Cancers 2022, 14, 4884. [Google Scholar] [CrossRef]
- Garrido, M.P.; Vega, M.; Romero, C. Antitumoral Effects of Metformin in Ovarian Cancer. Metformin 2019, 10, 163–180. [Google Scholar] [CrossRef]
- Drzewoski, J.; Hanefeld, M. The Current and Potential Therapeutic Use of Metformin-The Good Old Drug. Pharmaceuticals 2021, 14, 122. [Google Scholar] [CrossRef]
- Lashen, H. Role of metformin in the management of polycystic ovary syndrome. Ther. Adv. Endocrinol. Metab. 2010, 1, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Goswami, S.; Giacomini, K.M.; Altman, R.B.; Klein, T.E. Metformin pathways: Pharmacokinetics and pharmacodynamics. Pharm. Genom. 2012, 22, 820–827. [Google Scholar] [CrossRef]
- Motohashi, H.; Inui, K. Organic cation transporter OCTs (SLC22) and MATEs (SLC47) in the human kidney. AAPS J. 2013, 15, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Ota, K.; Ito, K.; Akahira, J.; Sato, N.; Onogawa, T.; Moriya, T.; Unno, M.; Abe, T.; Niikura, H.; Takano, T.; et al. Expression of organic cation transporter SLC22A16 in human epithelial ovarian cancer: A possible role of the adriamycin importer. Int. J. Gynecol. Pathol. 2007, 26, 334–340. [Google Scholar] [CrossRef]
- Andreev, E.; Brosseau, N.; Carmona, E.; Mes-Masson, A.M.; Ramotar, D. The human organic cation transporter OCT1 mediates high affinity uptake of the anticancer drug daunorubicin. Sci. Rep. 2016, 6, 20508. [Google Scholar] [CrossRef] [PubMed]
- Checkley, L.A.; Rudolph, M.C.; Wellberg, E.A.; Giles, E.D.; Wahdan-Alaswad, R.S.; Houck, J.A.; Edgerton, S.M.; Thor, A.D.; Schedin, P.; Anderson, S.M.; et al. Metformin Accumulation Correlates with Organic Cation Transporter 2 Protein Expression and Predicts Mammary Tumor Regression In Vivo. Cancer Prev. Res. 2017, 10, 198–207. [Google Scholar] [CrossRef]
- Bridgeman, S.C.; Ellison, G.C.; Melton, P.E.; Newsholme, P.; Mamotte, C.D.S. Epigenetic effects of metformin: From molecular mechanisms to clinical implications. Diabetes Obes. Metab. 2018, 20, 1553–1562. [Google Scholar] [CrossRef]
- Lengyel, E.; Litchfield, L.M.; Mitra, A.K.; Nieman, K.M.; Mukherjee, A.; Zhang, Y.; Johnson, A.; Bradaric, M.; Lee, W.; Romero, I.L. Metformin inhibits ovarian cancer growth and increases sensitivity to paclitaxel in mouse models. Am. J. Obstet. Gynecol. 2015, 212, 479.e1–479.e10. [Google Scholar] [CrossRef]
- Garrido, M.P.; Vera, C.; Vega, M.; Quest, A.F.G.; Romero, C. Metformin prevents nerve growth factor-dependent proliferative and proangiogenic effects in epithelial ovarian cancer cells and endothelial cells. Ther. Adv. Med. Oncol. 2018, 10, 1758835918770984. [Google Scholar] [CrossRef]
- Rattan, R.; Graham, R.P.; Maguire, J.L.; Giri, S.; Shridhar, V. Metformin suppresses ovarian cancer growth and metastasis with enhancement of cisplatin cytotoxicity in vivo. Neoplasia 2011, 13, 483–491. [Google Scholar] [CrossRef]
- Wu, B.; Li, S.; Sheng, L.; Zhu, J.; Gu, L.; Shen, H.; La, D.; Hambly, B.D.; Bao, S.; Di, W. Metformin inhibits the development and metastasis of ovarian cancer. Oncol. Rep. 2012, 28, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.; Plas, D.R.; Kubek, S.; Buzzai, M.; Mu, J.; Xu, Y.; Birnbaum, M.J.; Thompson, C.B. AMP-activated protein kinase induces a p53-dependent metabolic checkpoint. Mol. Cell 2005, 18, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Wei, J.; Wan, J.; Wang, W.; Wang, L.; Yuan, Y.; Yang, Z.; Liu, X.; Ming, L. Low glucose and metformin-induced apoptosis of human ovarian cancer cells is connected to ASK1 via mitochondrial and endoplasmic reticulum stress-associated pathways. J. Exp. Clin. Cancer Res. 2019, 38, 77. [Google Scholar] [CrossRef] [PubMed]
- Soraya, H.; Esfahanian, N.; Shakiba, Y.; Ghazi-Khansari, M.; Nikbin, B.; Hafezzadeh, H.; Maleki Dizaji, N.; Garjani, A. Anti-angiogenic Effects of Metformin, an AMPK Activator, on Human Umbilical Vein Endothelial Cells and on Granulation Tissue in Rat. Iran. J. Basic Med. Sci. 2012, 15, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C.; Sun, X.; Ma, Q.; Fu, G.F.; Cong, L.L.; Zhang, H.; Fan, D.F.; Feng, J.; Lu, S.Y.; Liu, J.L.; et al. Metformin’s antitumour and anti-angiogenic activities are mediated by skewing macrophage polarization. J. Cell. Mol. Med. 2018, 22, 3825–3836. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, P.; Wang, H.; Hou, D.; Li, W.; Xiao, G.; Li, C. Inhibitory effects of metformin at low concentration on epithelial-mesenchymal transition of CD44(+)CD117(+) ovarian cancer stem cells. Stem Cell Res. Ther. 2015, 6, 262. [Google Scholar] [CrossRef]
- Grassi, M.L.; Palma, C.S.; Thome, C.H.; Lanfredi, G.P.; Poersch, A.; Faca, V.M. Proteomic analysis of ovarian cancer cells during epithelial-mesenchymal transition (EMT) induced by epidermal growth factor (EGF) reveals mechanisms of cell cycle control. J. Proteom. 2017, 151, 2–11. [Google Scholar] [CrossRef]
- Chowanadisai, W.; Messerli, S.M.; Miller, D.H.; Medina, J.E.; Hamilton, J.W.; Messerli, M.A.; Brodsky, A.S. Cisplatin Resistant Spheroids Model Clinically Relevant Survival Mechanisms in Ovarian Tumors. PLoS ONE 2016, 11, e0151089. [Google Scholar] [CrossRef]
- Haslehurst, A.M.; Koti, M.; Dharsee, M.; Nuin, P.; Evans, K.; Geraci, J.; Childs, T.; Chen, J.; Li, J.; Weberpals, J.; et al. EMT transcription factors snail and slug directly contribute to cisplatin resistance in ovarian cancer. BMC Cancer 2012, 12, 91. [Google Scholar] [CrossRef]
- Kajiyama, H.; Shibata, K.; Terauchi, M.; Yamashita, M.; Ino, K.; Nawa, A.; Kikkawa, F. Chemoresistance to paclitaxel induces epithelial-mesenchymal transition and enhances metastatic potential for epithelial ovarian carcinoma cells. Int. J. Oncol. 2007, 31, 277–283. [Google Scholar] [CrossRef]
- Gonzalez, V.D.; Samusik, N.; Chen, T.J.; Savig, E.S.; Aghaeepour, N.; Quigley, D.A.; Huang, Y.W.; Giangarra, V.; Borowsky, A.D.; Hubbard, N.E.; et al. Commonly Occurring Cell Subsets in High-Grade Serous Ovarian Tumors Identified by Single-Cell Mass Cytometry. Cell Rep. 2018, 22, 1875–1888. [Google Scholar] [CrossRef]
- Zhu, X.; Shen, H.; Yin, X.; Long, L.; Xie, C.; Liu, Y.; Hui, L.; Lin, X.; Fang, Y.; Cao, Y.; et al. miR-186 regulation of Twist1 and ovarian cancer sensitivity to cisplatin. Oncogene 2016, 35, 323–332. [Google Scholar] [CrossRef]
- Cui, Y.; Qin, L.; Tian, D.; Wang, T.; Fan, L.; Zhang, P.; Wang, Z. ZEB1 Promotes Chemoresistance to Cisplatin in Ovarian Cancer Cells by Suppressing SLC3A2. Chemotherapy 2018, 63, 262–271. [Google Scholar] [CrossRef]
- Brown, J.R.; Chan, D.K.; Shank, J.J.; Griffith, K.A.; Fan, H.; Szulawski, R.; Yang, K.; Reynolds, R.K.; Johnston, C.; McLean, K.; et al. Phase II clinical trial of metformin as a cancer stem cell-targeting agent in ovarian cancer. JCI Insight 2020, 5, e133247. [Google Scholar] [CrossRef]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Hock, J.; Meister, G. The Argonaute protein family. Genome Biol. 2008, 9, 210. [Google Scholar] [CrossRef]
- Iwakawa, H.O.; Tomari, Y. Life of RISC: Formation, action, and degradation of RNA-induced silencing complex. Mol. Cell 2022, 82, 30–43. [Google Scholar] [CrossRef]
- Schulten, H.J. Pleiotropic Effects of Metformin on Cancer. Int. J. Mol. Sci. 2018, 19, 2850. [Google Scholar] [CrossRef]
- Alimoradi, N.; Firouzabadi, N.; Fatehi, R. How metformin affects various malignancies by means of microRNAs: A brief review. Cancer Cell Int. 2021, 21, 207. [Google Scholar] [CrossRef]
- Sargolzaei, J.; Etemadi, T.; Alyasin, A. The P53/microRNA network: A potential tumor suppressor with a role in anticancer therapy. Pharmacol. Res. 2020, 160, 105179. [Google Scholar] [CrossRef]
- Do, M.T.; Kim, H.G.; Choi, J.H.; Jeong, H.G. Metformin induces microRNA-34a to downregulate the Sirt1/Pgc-1alpha/Nrf2 pathway, leading to increased susceptibility of wild-type p53 cancer cells to oxidative stress and therapeutic agents. Free Radic. Biol. Med. 2014, 74, 21–34. [Google Scholar] [CrossRef]
- Sun, R.; Ma, X.; Cai, X.; Pan, X.; Liu, D. The effect and mechanism of action of metformin on in vitro FaDu cell proliferation. J. Int. Med. Res. 2016, 44, 1049–1054. [Google Scholar] [CrossRef]
- Sachdeva, M.; Zhu, S.; Wu, F.; Wu, H.; Walia, V.; Kumar, S.; Elble, R.; Watabe, K.; Mo, Y.Y. p53 represses c-Myc through induction of the tumor suppressor miR-145. Proc. Natl. Acad. Sci. USA 2009, 106, 3207–3212. [Google Scholar] [CrossRef]
- Suzuki, H.I.; Yamagata, K.; Sugimoto, K.; Iwamoto, T.; Kato, S.; Miyazono, K. Modulation of microRNA processing by p53. Nature 2009, 460, 529–533. [Google Scholar] [CrossRef]
- Beaufort, C.M.; Helmijr, J.C.; Piskorz, A.M.; Hoogstraat, M.; Ruigrok-Ritstier, K.; Besselink, N.; Murtaza, M.; van IJcken, W.F.J.; Heine, A.A.; Smid, M.; et al. Ovarian cancer cell line panel (OCCP): Clinical importance of in vitro morphological subtypes. PLoS ONE 2014, 9, e103988. [Google Scholar] [CrossRef]
- Li, C.; Liu, V.W.; Chan, D.W.; Yao, K.M.; Ngan, H.Y. LY294002 and metformin cooperatively enhance the inhibition of growth and the induction of apoptosis of ovarian cancer cells. Int. J. Gynecol. Cancer 2012, 22, 15–22. [Google Scholar] [CrossRef]
- Dong, R.; Liu, X.; Zhang, Q.; Jiang, Z.; Li, Y.; Wei, Y.; Li, Y.; Yang, Q.; Liu, J.; Wei, J.J.; et al. miR-145 inhibits tumor growth and metastasis by targeting metadherin in high-grade serous ovarian carcinoma. Oncotarget 2014, 5, 10816–10829. [Google Scholar] [CrossRef]
- Rechsteiner, M.; Zimmermann, A.K.; Wild, P.J.; Caduff, R.; von Teichman, A.; Fink, D.; Moch, H.; Noske, A. TP53 mutations are common in all subtypes of epithelial ovarian cancer and occur concomitantly with KRAS mutations in the mucinous type. Exp. Mol. Pathol. 2013, 95, 235–241. [Google Scholar] [CrossRef]
- Gralewska, P.; Gajek, A.; Marczak, A.; Rogalska, A. Metformin Affects Olaparib Sensitivity through Induction of Apoptosis in Epithelial Ovarian Cancer Cell Lines. Int. J. Mol. Sci. 2021, 22, 557. [Google Scholar] [CrossRef]
- Sun, Y.; Tao, C.; Huang, X.; He, H.; Shi, H.; Zhang, Q.; Wu, H. Metformin induces apoptosis of human hepatocellular carcinoma HepG2 cells by activating an AMPK/p53/miR-23a/FOXA1 pathway. OncoTargets Ther. 2016, 9, 2845–2853. [Google Scholar] [CrossRef]
- Chen, H.Z.; Tsai, S.Y.; Leone, G. Emerging roles of E2Fs in cancer: An exit from cell cycle control. Nat. Rev. Cancer 2009, 9, 785–797. [Google Scholar] [CrossRef]
- Gao, Y.; Li, H.; Ma, X.; Fan, Y.; Ni, D.; Zhang, Y.; Huang, Q.; Liu, K.; Li, X.; Wang, L.; et al. E2F3 upregulation promotes tumor malignancy through the transcriptional activation of HIF-2alpha in clear cell renal cell carcinoma. Oncotarget 2017, 8, 54021–54036. [Google Scholar] [CrossRef]
- Pulito, C.; Mori, F.; Sacconi, A.; Goeman, F.; Ferraiuolo, M.; Pasanisi, P.; Campagnoli, C.; Berrino, F.; Fanciulli, M.; Ford, R.J.; et al. Metformin-induced ablation of microRNA 21-5p releases Sestrin-1 and CAB39L antitumoral activities. Cell Discov. 2017, 3, 17022. [Google Scholar] [CrossRef]
- Blandino, G.; Valerio, M.; Cioce, M.; Mori, F.; Casadei, L.; Pulito, C.; Sacconi, A.; Biagioni, F.; Cortese, G.; Galanti, S.; et al. Metformin elicits anticancer effects through the sequential modulation of DICER and c-MYC. Nat. Commun. 2012, 3, 865. [Google Scholar] [CrossRef]
- Reimer, D.; Hubalek, M.; Kiefel, H.; Riedle, S.; Skvortsov, S.; Erdel, M.; Hofstetter, G.; Concin, N.; Fiegl, H.; Muller-Holzner, E.; et al. Regulation of transcription factor E2F3a and its clinical relevance in ovarian cancer. Oncogene 2011, 30, 4038–4049. [Google Scholar] [CrossRef]
- Garrido, M.P.; Torres, I.; Avila, A.; Chnaiderman, J.; Valenzuela-Valderrama, M.; Aramburo, J.; Orostica, L.; Duran-Jara, E.; Lobos-Gonzalez, L.; Romero, C. NGF/TRKA Decrease miR-145-5p Levels in Epithelial Ovarian Cancer Cells. Int. J. Mol. Sci. 2020, 21, 7657. [Google Scholar] [CrossRef]
- Hua, M.; Qin, Y.; Sheng, M.; Cui, X.; Chen, W.; Zhong, J.; Yan, J.; Chen, Y. miR-145 suppresses ovarian cancer progression via modulation of cell growth and invasion by targeting CCND2 and E2F3. Mol. Med. Rep. 2019, 19, 3575–3583. [Google Scholar] [CrossRef]
- Kanwal, R.; Gupta, S. Epigenetic modifications in cancer. Clin. Genet. 2012, 81, 303–311. [Google Scholar] [CrossRef]
- Yan, L.; Zhou, J.; Gao, Y.; Ghazal, S.; Lu, L.; Bellone, S.; Yang, Y.; Liu, N.; Zhao, X.; Santin, A.D.; et al. Regulation of tumor cell migration and invasion by the H19/let-7 axis is antagonized by metformin-induced DNA methylation. Oncogene 2015, 34, 3076–3084. [Google Scholar] [CrossRef]
- Kallen, A.N.; Zhou, X.B.; Xu, J.; Qiao, C.; Ma, J.; Yan, L.; Lu, L.; Liu, C.; Yi, J.S.; Zhang, H.; et al. The imprinted H19 lncRNA antagonizes let-7 microRNAs. Mol. Cell 2013, 52, 101–112. [Google Scholar] [CrossRef]
- Chirshev, E.; Hojo, N.; Bertucci, A.; Sanderman, L.; Nguyen, A.; Wang, H.; Suzuki, T.; Brito, E.; Martinez, S.R.; Castanon, C.; et al. Epithelial/mesenchymal heterogeneity of high-grade serous ovarian carcinoma samples correlates with miRNA let-7 levels and predicts tumor growth and metastasis. Mol. Oncol. 2020, 14, 2796–2813. [Google Scholar] [CrossRef]
- Wu, P.; Tang, Y.; Fang, X.; Xie, C.; Zeng, J.; Wang, W.; Zhao, S. Metformin Suppresses Hypopharyngeal Cancer Growth by Epigenetically Silencing Long Non-coding RNA SNHG7 in FaDu Cells. Front. Pharmacol. 2019, 10, 143. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, Y.; Wang, B.; Zhai, J. Metformin Affects Paclitaxel Sensitivity of Ovarian Cancer Cells Through Autophagy Mediated by Long Noncoding RNASNHG7/miR-3127-5p Axis. Cancer Biother. Radiopharm. 2022, 37, 792–801. [Google Scholar] [CrossRef]
- Gu, Y.; Zhang, B.; Gu, G.; Yang, X.; Qian, Z. Metformin Increases the Chemosensitivity of Pancreatic Cancer Cells to Gemcitabine by Reversing EMT Through Regulation DNA Methylation of miR-663. OncoTargets Ther. 2020, 13, 10417–10429. [Google Scholar] [CrossRef]
- Bao, X.; Zhao, L.; Guan, H.; Li, F. Inhibition of LCMR1 and ATG12 by demethylation-activated miR-570-3p is involved in the anti-metastasis effects of metformin on human osteosarcoma. Cell Death Dis. 2018, 9, 611. [Google Scholar] [CrossRef]
- Li, J.; Lu, J.; Ye, Z.; Han, X.; Zheng, X.; Hou, H.; Chen, W.; Li, X.; Zhao, L. 20(S)-Rg3 blocked epithelial-mesenchymal transition through DNMT3A/miR-145/FSCN1 in ovarian cancer. Oncotarget 2017, 8, 53375–53386. [Google Scholar] [CrossRef]
- Wu, H.; Xiao, Z.; Wang, K.; Liu, W.; Hao, Q. MiR-145 is downregulated in human ovarian cancer and modulates cell growth and invasion by targeting p70S6K1 and MUC1. Biochem. Biophys. Res. Commun. 2013, 441, 693–700. [Google Scholar] [CrossRef]
- Garrido, M.P.; Salvatierra, R.; Valenzuela-Valderrama, M.; Vallejos, C.; Bruneau, N.; Hernandez, A.; Vega, M.; Selman, A.; Quest, A.F.G.; Romero, C. Metformin Reduces NGF-Induced Tumour Promoter Effects in Epithelial Ovarian Cancer Cells. Pharmaceuticals 2020, 13, 315. [Google Scholar] [CrossRef]
- Cuyas, E.; Fernandez-Arroyo, S.; Verdura, S.; Garcia, R.A.; Stursa, J.; Werner, L.; Blanco-Gonzalez, E.; Montes-Bayon, M.; Joven, J.; Viollet, B.; et al. Metformin regulates global DNA methylation via mitochondrial one-carbon metabolism. Oncogene 2018, 37, 963–970. [Google Scholar] [CrossRef]
- Zhong, T.; Men, Y.; Lu, L.; Geng, T.; Zhou, J.; Mitsuhashi, A.; Shozu, M.; Maihle, N.J.; Carmichael, G.G.; Taylor, H.S.; et al. Metformin alters DNA methylation genome-wide via the H19/SAHH axis. Oncogene 2017, 36, 2345–2354. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.B.; Kim, D.; Kim, Y.; Han, J.; Shim, Y.M.; Kim, D.H. Metformin regulates expression of DNA methyltransferases through the miR-148/-152 family in non-small lung cancer cells. Clin. Epigenetics 2023, 15, 48. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Chen, C.; Ji, X.; Liu, J.; Zhou, Q.; Wang, G.; Yuan, W.; Kan, Q.; Sun, Z. The interplay between m6A RNA methylation and noncoding RNA in cancer. J. Hematol. Oncol. 2019, 12, 121. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Gao, S.; Ma, L.; Sun, Y.; Peng, Z.Y.; Wu, J.; Du, N.; Ren, H.; Tang, S.C.; Sun, X. Stimulation of Let-7 Maturation by Metformin Improved the Response to Tyrosine Kinase Inhibitor Therapy in an m6A Dependent Manner. Front. Oncol. 2021, 11, 731561. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Guan, H.; Lin, X.; Li, N.; Geng, F.; Li, J. METTL3 serves an oncogenic role in human ovarian cancer cells partially via the AKT signaling pathway. Oncol. Lett. 2020, 19, 3197–3204. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, X.; Huang, Y.Z.; Zhu, Y.L.; Xu, L.Y.; Li, Z.; Dai, X.Y.; Shi, L.; Zhou, X.J.; Wei, J.F.; et al. Metformin exhibits antiproliferation activity in breast cancer via miR-483-3p/METTL3/m(6)A/p21 pathway. Oncogenesis 2021, 10, 7. [Google Scholar] [CrossRef]
- Gurtner, A.; Falcone, E.; Garibaldi, F.; Piaggio, G. Dysregulation of microRNA biogenesis in cancer: The impact of mutant p53 on Drosha complex activity. J. Exp. Clin. Cancer Res. 2016, 35, 45. [Google Scholar] [CrossRef]
- Thomson, J.M.; Newman, M.; Parker, J.S.; Morin-Kensicki, E.M.; Wright, T.; Hammond, S.M. Extensive post-transcriptional regulation of microRNAs and its implications for cancer. Genes Dev. 2006, 20, 2202–2207. [Google Scholar] [CrossRef]
- Jiang, F.Z.; He, Y.Y.; Wang, H.H.; Zhang, H.L.; Zhang, J.; Yan, X.F.; Wang, X.J.; Che, Q.; Ke, J.Q.; Chen, Z.; et al. Mutant p53 induces EZH2 expression and promotes epithelial-mesenchymal transition by disrupting p68-Drosha complex assembly and attenuating miR-26a processing. Oncotarget 2015, 6, 44660–44674. [Google Scholar] [CrossRef]
- Merritt, W.M.; Lin, Y.G.; Han, L.Y.; Kamat, A.A.; Spannuth, W.A.; Schmandt, R.; Urbauer, D.; Pennacchio, L.A.; Cheng, J.F.; Nick, A.M.; et al. Dicer, Drosha, and outcomes in patients with ovarian cancer. N. Engl. J. Med. 2008, 359, 2641–2650. [Google Scholar] [CrossRef]
- Kuang, Y.; Cai, J.; Li, D.; Han, Q.; Cao, J.; Wang, Z. Repression of Dicer is associated with invasive phenotype and chemoresistance in ovarian cancer. Oncol. Lett. 2013, 5, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Cai, L.; Guo, J.; Chen, N.; Yi, X.; Zhao, Y.; Cai, J.; Wang, Z. Depletion of Dicer promotes epithelial ovarian cancer progression by elevating PDIA3 expression. Tumour Biol. 2016, 37, 14009–14023. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.S.; Ip, C.K.; Mak, A.S.; Wong, A.S. A novel p70 S6 kinase-microRNA biogenesis axis mediates multicellular spheroid formation in ovarian cancer progression. Oncotarget 2016, 7, 38064–38077. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.L.; Zhang, Q.H.; Wang, X.W.; He, H. Antidiabetic drug metformin mitigates ovarian cancer SKOV3 cell growth by triggering G2/M cell cycle arrest and inhibition of m-TOR/PI3K/Akt signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1169–1175. [Google Scholar] [PubMed]
- Gwak, H.; Kim, Y.; An, H.; Dhanasekaran, D.N.; Song, Y.S. Metformin induces degradation of cyclin D1 via AMPK/GSK3beta axis in ovarian cancer. Mol. Carcinog. 2017, 56, 349–358. [Google Scholar] [CrossRef]
- Retamales-Ortega, R.; Orostica, L.; Vera, C.; Cuevas, P.; Hernandez, A.; Hurtado, I.; Vega, M.; Romero, C. Role of Nerve Growth Factor (NGF) and miRNAs in Epithelial Ovarian Cancer. Int. J. Mol. Sci. 2017, 18, 507. [Google Scholar] [CrossRef]
- Gao, P.; Tchernyshyov, I.; Chang, T.C.; Lee, Y.S.; Kita, K.; Ochi, T.; Zeller, K.I.; De Marzo, A.M.; Van Eyk, J.E.; Mendell, J.T.; et al. c-Myc suppression of miR-23a/b enhances mitochondrial glutaminase expression and glutamine metabolism. Nature 2009, 458, 762–765. [Google Scholar] [CrossRef]
- Zeinali, T.; Mansoori, B.; Mohammadi, A.; Baradaran, B. Regulatory mechanisms of miR-145 expression and the importance of its function in cancer metastasis. Biomed. Pharmacother. 2019, 109, 195–207. [Google Scholar] [CrossRef]
- Hao, B.; Zhang, J. miRNA-21 inhibition suppresses the human epithelial ovarian cancer by targeting PTEN signal pathway. Saudi J. Biol. Sci. 2019, 26, 2026–2029. [Google Scholar] [CrossRef]
- Liu, H.Y.; Zhang, Y.Y.; Zhu, B.L.; Feng, F.Z.; Yan, H.; Zhang, H.Y.; Zhou, B. miR-21 regulates the proliferation and apoptosis of ovarian cancer cells through PTEN/PI3K/AKT. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4149–4155. [Google Scholar] [CrossRef]
- Liu, S.; Fang, Y.; Shen, H.; Xu, W.; Li, H. Berberine sensitizes ovarian cancer cells to cisplatin through miR-21/PDCD4 axis. Acta Biochim. Biophys. Sin. 2013, 45, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Yang, X.; Wang, F.; Cui, Z.; Huang, Y. MicroRNA-21 promotes the cell proliferation, invasion and migration abilities in ovarian epithelial carcinomas through inhibiting the expression of PTEN protein. Int. J. Mol. Med. 2010, 26, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Huo, W.; Zhao, G.; Yin, J.; Ouyang, X.; Wang, Y.; Yang, C.; Wang, B.; Dong, P.; Wang, Z.; Watari, H.; et al. Lentiviral CRISPR/Cas9 vector mediated miR-21 gene editing inhibits the epithelial to mesenchymal transition in ovarian cancer cells. J. Cancer 2017, 8, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Kumar, S. Metformin inhibits human breast cancer cell growth by promoting apoptosis via a ROS-independent pathway involving mitochondrial dysfunction: Pivotal role of superoxide dismutase (SOD). Cell. Oncol. 2018, 41, 637–650. [Google Scholar] [CrossRef]
- Nangia-Makker, P.; Yu, Y.; Vasudevan, A.; Farhana, L.; Rajendra, S.G.; Levi, E.; Majumdar, A.P. Metformin: A potential therapeutic agent for recurrent colon cancer. PLoS ONE 2014, 9, e84369. [Google Scholar] [CrossRef]
- Kalogirou, C.; Schafer, D.; Krebs, M.; Kurz, F.; Schneider, A.; Riedmiller, H.; Kneitz, B.; Vergho, D. Metformin-Derived Growth Inhibition in Renal Cell Carcinoma Depends on miR-21-Mediated PTEN Expression. Urol. Int. 2016, 96, 106–115. [Google Scholar] [CrossRef]
- Tossetta, G. Metformin Improves Ovarian Cancer Sensitivity to Paclitaxel and Platinum-Based Drugs: A Review of In Vitro Findings. Int. J. Mol. Sci. 2022, 23, 2893. [Google Scholar] [CrossRef]
- Luo, M.; Tan, X.; Mu, L.; Luo, Y.; Li, R.; Deng, X.; Chen, N.; Ren, M.; Li, Y.; Wang, L.; et al. MiRNA-21 mediates the antiangiogenic activity of metformin through targeting PTEN and SMAD7 expression and PI3K/AKT pathway. Sci. Rep. 2017, 7, 43427. [Google Scholar] [CrossRef]
- Echevarria-Vargas, I.M.; Valiyeva, F.; Vivas-Mejia, P.E. Upregulation of miR-21 in cisplatin resistant ovarian cancer via JNK-1/c-Jun pathway. PLoS ONE 2014, 9, e97094. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, X.; Liu, J.; Sun, B.; Tang, H.; Zhang, H. miR-27a-mediated antiproliferative effects of metformin on the breast cancer cell line MCF-7. Oncol. Rep. 2016, 36, 3691–3699. [Google Scholar] [CrossRef]
- Nair, V.; Pathi, S.; Jutooru, I.; Sreevalsan, S.; Basha, R.; Abdelrahim, M.; Samudio, I.; Safe, S. Metformin inhibits pancreatic cancer cell and tumor growth and downregulates Sp transcription factors. Carcinogenesis 2013, 34, 2870–2879. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Sun, X.; Jiang, X. UCA1 involved in the metformin-regulated bladder cancer cell proliferation and glycolysis. Tumour Biol. 2017, 39, 1010428317710823. [Google Scholar] [CrossRef] [PubMed]
- Golshan, M.; Khaleghi, S.; Shafiee, S.M.; Valaee, S.; Ghanei, Z.; Jamshidizad, A.; Dashtizad, M.; Shamsara, M. Metformin modulates oncogenic expression of HOTAIR gene via promoter methylation and reverses epithelial-mesenchymal transition in MDA-MB-231 cells. J. Cell. Biochem. 2021, 122, 385–393. [Google Scholar] [CrossRef]
- Huang, Y.; Zhou, Z.; Zhang, J.; Hao, Z.; He, Y.; Wu, Z.; Song, Y.; Yuan, K.; Zheng, S.; Zhao, Q.; et al. lncRNA MALAT1 participates in metformin inhibiting the proliferation of breast cancer cell. J. Cell. Mol. Med. 2021, 25, 7135–7145. [Google Scholar] [CrossRef]
- Chen, J.; Qin, C.; Zhou, Y.; Chen, Y.; Mao, M.; Yang, J. Metformin may induce ferroptosis by inhibiting autophagy via lncRNA H19 in breast cancer. FEBS Open Bio 2022, 12, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Qian, T.; Li, S.; Xie, Y.; Tao, M. Metformin reverses tamoxifen resistance through the lncRNA GAS5-medicated mTOR pathway in breast cancer. Ann. Transl. Med. 2022, 10, 366. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, Y.; Chen, L.; Li, R.; Ning, Y.; Zhu, X. Role of metformin in functional endometrial hyperplasia and polycystic ovary syndrome involves the regulation of MEG3/miR-223/GLUT4 and SNHG20/miR-4486/GLUT4 signaling. Mol. Med. Rep. 2022, 26, 218. [Google Scholar] [CrossRef]
- Guo, J.; Li, Y.; Duan, H.; Yuan, L. Metformin Suppresses the Proliferation and Promotes the Apoptosis of Colon Cancer Cells Through Inhibiting the Expression of Long Noncoding RNA-UCA1. OncoTargets Ther. 2020, 13, 4169–4181. [Google Scholar] [CrossRef]
- Aminimoghaddam, S.; Fooladi, B.; Noori, M.; Nickhah Klashami, Z.; Kakavand Hamidi, A.; Amoli, M.M. The Effect of Metformin on Expression of Long Non-coding RNA H19 in Endometrial Cancer. Med. J. Islam Repub. Iran 2021, 35, 155. [Google Scholar] [CrossRef]
- Li, P.; Tong, L.; Song, Y.; Sun, J.; Shi, J.; Wu, Z.; Diao, Y.; Li, Y.; Wang, Z. Long noncoding RNA H19 participates in metformin-mediated inhibition of gastric cancer cell invasion. J. Cell. Physiol. 2019, 234, 4515–4527. [Google Scholar] [CrossRef]
- Tseng, H.H.; Chen, Y.Z.; Chou, N.H.; Chen, Y.C.; Wu, C.C.; Liu, L.F.; Yang, Y.F.; Yeh, C.Y.; Kung, M.L.; Tu, Y.T.; et al. Metformin inhibits gastric cancer cell proliferation by regulation of a novel Loc100506691-CHAC1 axis. Mol. Ther. Oncolytics 2021, 22, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Gandhy, S.U.; Imanirad, P.; Jin, U.H.; Nair, V.; Hedrick, E.; Cheng, Y.; Corton, J.C.; Kim, K.; Safe, S. Specificity protein (Sp) transcription factors and metformin regulate expression of the long non-coding RNA HULC. Oncotarget 2015, 6, 26359–26372. [Google Scholar] [CrossRef] [PubMed]
- Sabry, D.; Abdelaleem, O.O.; El Amin Ali, A.M.; Mohammed, R.A.; Abdel-Hameed, N.D.; Hassouna, A.; Khalifa, W.A. Anti-proliferative and anti-apoptotic potential effects of epigallocatechin-3-gallate and/or metformin on hepatocellular carcinoma cells: In vitro study. Mol. Biol. Rep. 2019, 46, 2039–2047. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Luo, M.; Zhang, J.; Guo, B.; Singh, S.; Lin, X.; Xiong, H.; Ju, S.; Wang, L.; Zhou, Y.; et al. The role of lncRNA H19 in tumorigenesis and drug resistance of human Cancers. Front. Genet 2022, 13, 1005522. [Google Scholar] [CrossRef]
- Ma, Y.; Yu, L.; Yan, W.; Qiu, L.; Zhang, J.; Jia, X. lncRNA GAS5 Sensitizes Breast Cancer Cells to Ionizing Radiation by Inhibiting DNA Repair. BioMed Res. Int. 2022, 2022, 1987519. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, W.J. Long non-coding RNA-H19 promotes ovarian cancer cell proliferation and migration via the microRNA-140/Wnt1 axis. Kaohsiung J. Med. Sci. 2021, 37, 768–775. [Google Scholar] [CrossRef]
- Mizrahi, A.; Czerniak, A.; Levy, T.; Amiur, S.; Gallula, J.; Matouk, I.; Abu-lail, R.; Sorin, V.; Birman, T.; de Groot, N.; et al. Development of targeted therapy for ovarian cancer mediated by a plasmid expressing diphtheria toxin under the control of H19 regulatory sequences. J. Transl. Med. 2009, 7, 69. [Google Scholar] [CrossRef]
- Ma, H.; Gao, L.; Yu, H.; Song, X. Long non-coding RNA H19 correlates with unfavorable prognosis and promotes cell migration and invasion in ovarian cancer. Ginekol. Pol. 2022, 93, 68293. [Google Scholar] [CrossRef]
- Zhao, L.; Sun, W.; Zheng, A.; Zhang, Y.; Fang, C.; Zhang, P. Ginsenoside Rg3 suppresses ovarian cancer cell proliferation and invasion by inhibiting the expression of lncRNA H19. Acta Biochim. Pol. 2021, 68, 575–582. [Google Scholar] [CrossRef]
- Zhu, Z.; Song, L.; He, J.; Sun, Y.; Liu, X.; Zou, X. Ectopic expressed long non-coding RNA H19 contributes to malignant cell behavior of ovarian cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 10082–10091. [Google Scholar]
- Xu, J.; Zheng, L.H.; Hong, Y.N.; Xuan, C.; Yan, S.L.; Lv, G.L.; Jiang, Z.G.; Ding, X.F. Long Non-coding RNA UCA1 Regulates SRPK1 Expression Through miR- 99b-3p in Ovarian Cancer. Protein Pept. Lett. 2022, 29, 829–838. [Google Scholar] [CrossRef]
- Zheng, X.; Zhou, Y.; Chen, W.; Chen, L.; Lu, J.; He, F.; Li, X.; Zhao, L. Ginsenoside 20(S)-Rg3 Prevents PKM2-Targeting miR-324-5p from H19 Sponging to Antagonize the Warburg Effect in Ovarian Cancer Cells. Cell Physiol. Biochem. 2018, 51, 1340–1353. [Google Scholar] [CrossRef]
- Tian, X.; Zuo, X.; Hou, M.; Li, C.; Teng, Y. LncRNA-H19 regulates chemoresistance to carboplatin in epithelial ovarian cancer through microRNA-29b-3p and STAT3. J. Cancer 2021, 12, 5712–5722. [Google Scholar] [CrossRef]
- Hashemi, M.; Moosavi, M.S.; Abed, H.M.; Dehghani, M.; Aalipour, M.; Heydari, E.A.; Behroozaghdam, M.; Entezari, M.; Salimimoghadam, S.; Gunduz, E.S.; et al. Long non-coding RNA (lncRNA) H19 in human cancer: From proliferation and metastasis to therapy. Pharmacol. Res. 2022, 184, 106418. [Google Scholar] [CrossRef]
- Zheng, Z.G.; Xu, H.; Suo, S.S.; Xu, X.L.; Ni, M.W.; Gu, L.H.; Chen, W.; Wang, L.Y.; Zhao, Y.; Tian, B.; et al. The Essential Role of H19 Contributing to Cisplatin Resistance by Regulating Glutathione Metabolism in High-Grade Serous Ovarian Cancer. Sci. Rep. 2016, 6, 26093. [Google Scholar] [CrossRef] [PubMed]
- Medrzycki, M.; Zhang, Y.; Zhang, W.; Cao, K.; Pan, C.; Lailler, N.; McDonald, J.F.; Bouhassira, E.E.; Fan, Y. Histone h1.3 suppresses h19 noncoding RNA expression and cell growth of ovarian cancer cells. Cancer Res. 2014, 74, 6463–6473. [Google Scholar] [CrossRef] [PubMed]
- Malakoti, F.; Alemi, F.; Yeganeh, S.J.; Hosseini, F.; Shabestani, N.; Samemaleki, S.; Maleki, M.; Daneshvar, S.F.; Montazer, M.; Yousefi, B. Long noncoding RNA SNHG7-miRNA-mRNA axes crosstalk with oncogenic signaling pathways in human cancers. Chem. Biol. Drug Des. 2023, 101, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Wu, Y.; Bai, S.; Yan, Y.; Kang, H.; Ma, W.; Zhang, J.; Gao, Y.; Hui, B.; Ma, H.; et al. Long non-coding RNA SNGH7 Is activated by SP1 and exerts oncogenic properties by interacting with EZH2 in ovarian cancer. J. Cell. Mol. Med. 2020, 24, 7479–7489. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, R.; Ye, Y. Long non-coding RNA (LncRNA) SNHG7/Eukaryotic translation initiation factor 4 gamma 2 (EIF4G2) involves in the malignant events of ovarian cancer cells with paclitaxel resistant. Bioengineered 2021, 12, 10541–10552. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, Y.; He, J.; Sun, H.; Jin, Z. Long non-coding RNA H19 mediates ovarian cancer cell cisplatin-resistance and migration during EMT. Int. J. Clin. Exp. Pathol. 2019, 12, 2506–2515. [Google Scholar]
- Goodwin, P.J.; Chen, B.E.; Gelmon, K.A.; Whelan, T.J.; Ennis, M.; Lemieux, J.; Ligibel, J.A.; Hershman, D.L.; Mayer, I.A.; Hobday, T.J.; et al. Effect of Metformin vs. Placebo on Invasive Disease-Free Survival in Patients with Breast Cancer: The MA.32 Randomized Clinical Trial. JAMA 2022, 327, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- Kordes, S.; Pollak, M.N.; Zwinderman, A.H.; Mathot, R.A.; Weterman, M.J.; Beeker, A.; Punt, C.J.; Richel, D.J.; Wilmink, J.W. Metformin in patients with advanced pancreatic cancer: A double-blind, randomised, placebo-controlled phase 2 trial. Lancet Oncol. 2015, 16, 839–847. [Google Scholar] [CrossRef]
- Lord, S.R.; Cheng, W.C.; Liu, D.; Gaude, E.; Haider, S.; Metcalf, T.; Patel, N.; Teoh, E.J.; Gleeson, F.; Bradley, K.; et al. Integrated Pharmacodynamic Analysis Identifies Two Metabolic Adaption Pathways to Metformin in Breast Cancer. Cell Metab. 2018, 28, 679–688.e674. [Google Scholar] [CrossRef] [PubMed]
- Higurashi, T.; Hosono, K.; Takahashi, H.; Komiya, Y.; Umezawa, S.; Sakai, E.; Uchiyama, T.; Taniguchi, L.; Hata, Y.; Uchiyama, S.; et al. Metformin for chemoprevention of metachronous colorectal adenoma or polyps in post-polypectomy patients without diabetes: A multicentre double-blind, placebo-controlled, randomised phase 3 trial. Lancet Oncol. 2016, 17, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Gutkind, J.S.; Molinolo, A.A.; Wu, X.; Wang, Z.; Nachmanson, D.; Harismendy, O.; Alexandrov, L.B.; Wuertz, B.R.; Ondrey, F.G.; Laronde, D.; et al. Inhibition of mTOR signaling and clinical activity of metformin in oral premalignant lesions. JCI Insight 2021, 6, e147096. [Google Scholar] [CrossRef] [PubMed]
- Laskov, I.; Drudi, L.; Beauchamp, M.C.; Yasmeen, A.; Ferenczy, A.; Pollak, M.; Gotlieb, W.H. Anti-diabetic doses of metformin decrease proliferation markers in tumors of patients with endometrial cancer. Gynecol. Oncol. 2014, 134, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Hadad, S.; Iwamoto, T.; Jordan, L.; Purdie, C.; Bray, S.; Baker, L.; Jellema, G.; Deharo, S.; Hardie, D.G.; Pusztai, L.; et al. Evidence for biological effects of metformin in operable breast cancer: A pre-operative, window-of-opportunity, randomized trial. Breast Cancer Res. Treat. 2011, 128, 783–794. [Google Scholar] [CrossRef]
- Joshua, A.M.; Zannella, V.E.; Downes, M.R.; Bowes, B.; Hersey, K.; Koritzinsky, M.; Schwab, M.; Hofmann, U.; Evans, A.; van der Kwast, T.; et al. A pilot ‘window of opportunity’ neoadjuvant study of metformin in localised prostate cancer. Prostate Cancer Prostatic Dis. 2014, 17, 252–258. [Google Scholar] [CrossRef]
- Mitsuhashi, A.; Kiyokawa, T.; Sato, Y.; Shozu, M. Effects of metformin on endometrial cancer cell growth in vivo: A preoperative prospective trial. Cancer 2014, 120, 2986–2995. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Evaluation of Metformin Effect on the Fertility of Women Treated With 131I for Thyroid Cancer (METHYR). Available online: https://classic.clinicaltrials.gov/ct2/show/NCT05468554 (accessed on 23 August 2023).
- ClinicalTrials.gov. Cancer Chemoprevention by Metformin Hydrochloride in Oral Potentially Malignant Lesions. Available online: https://clinicaltrials.gov/study/NCT03685409 (accessed on 23 August 2023).
- ClinicalTrials.gov. Cancer Chemoprevention by Metformin Hydrochloride Compared to Placebo in Oral Potentially Malignant Lesions. Available online: https://clinicaltrials.gov/study/NCT03684707?id=%22NCT03684707%22&rank=1 (accessed on 23 August 2023).
- ClinicalTrials.gov. Longitudinal Follow-Up in Women with Endometrial Hyperplasia without Atypia. Available online: https://clinicaltrials.gov/study/NCT05292573?tab=results (accessed on 23 August 2023).