


1. Introduction
In dairy cattle herds, a large percentage of cows are affected by mastitis—inflammation of the mammary gland. The consequences of the occurrence of this disease are a decrease in animal welfare and a loss of animal performance due to decreased milk production and the necessity for veterinary treatment. Mastitis is mainly caused by bacteria [1], fungi, or algae [2]. To prevent mastitis, an appropriate hygiene standard (hygienic surroundings and disinfection of milking equipment) is crucial; however, due to the conditions in the barn, this is challenging to accomplish. These pathogens may originate from various sources, for instance, from the environment in which the animal is living [3]. Infection can occur through pathogens penetrating the teat canal and then entering the mammary gland, where inflammation may occur. The reservoir for these pathogens may be found not only in the surrounding environment but also on contaminated tools, other animals, or staff who do not follow hygiene procedures [1]. Pathogens that cause mastitis can be divided into environmental and infectious groups. Environmental pathogens include Escherichia coli, Klebsiella spp., and Candida spp., while the infectious group includes Staphylococcus aureus, Streptococcus agalactiae, Streptococcus dysgalactiae, Streptococcus uberis, and Mycoplasma bovis. High temperatures and humidity promote the development of various fungi that can induce mastitis, including Geotrichum spp., Trichosporon spp., Rhodotorula spp., and Torulopsis spp. However, the Candida genus’s yeasts are deemed to be the prevailing mycotic flora [4]. There are also known cases of mastitis induced by algae from the genus Prototheca [2].
The most common treatment for mastitis is antibiotic therapy; however, the long-term overuse and application of antibiotics in animal production as a form of prophylaxis has led to the development of resistant bacteria, further leading to the development of antibiotic resistance [5], which poses a threat to animal [6] and human [7] health. Herd management during the dry period, which is the period with no milk production, is crucial. At the same time, the dry period forms the transitional period between the last lactation, and calving and the next lactation; therefore, in order to prevent a recurrence of the disease, it is necessary to implement treatment to cure the inflammation and also to focus on preventing the occurrence of mastitis [8]. Due to the growing problem of antibiotic resistance, it is essential to look for alternatives that can be used in the treatment and prophylaxis of mastitis. The latest and most innovative solution for eliminating bacterial growth is the use of nanoparticles (NPs). These particles effectively damage the cell membranes of bacteria and the biofilms they are able to form [9], especially in the case of drug-resistant bacteria, such as Staphylococcus aureus, Staphylococcus epidermidis, and Escherichia coli [10].
The aim of this study was to conduct an in vitro analysis to determine the effect of nanoparticle (NP) hydrocolloids of AuNPs, CuNPs, AgNPs, PtNPs, iron nanoparticles with a surface carbon layer (NP-FeCs), and their triple complexes on the survival of the most common mastitis pathogens. Homogenous NPs were tested in various forms: (1) hydrocolloid, (2) mixed with beeswax, (3) mixed with olive oil, and (4) in a mixture based on vegetable glycerin and propylene glycol. The mentioned mixtures were used against the pathogens Staphylococcus aureus, Escherichia coli, Streptococcus agalactiae, Streptococcus uberis, Enterococcus faecalis, Enterobacter cloacae, Pseudomonas aeruginosa, and Candida albicans. The effect of the NPs in hydrocolloid and mixture forms on the viability of BME-UV1 epithelial mammary cells was also determined.
2. Materials and Methods
2.1. Preparation of Homogenous Metal NPs and Their Complexes Using the Self-Organization Phenomenon
Solutions of NP complexes were prepared by mixing AgNP, CuNP, and AuNP hydrocolloids at a concentration of 50 mg/L (Nano-Tech, Warsaw, Poland) and NP-FeC hydrocolloids at a concentration of 50 mg/L (PlasmaChem, Berlin, Germany) at a 1:1 ratio. All the prepared NP solutions were incubated for 24 h at a temperature of 24 °C and then used for further analysis. Homogenous hydrocolloids were stored at room temperature. The NPs from Nano-Tech and the NP-FeCs from PlasmaChem were synthesized using physical methods. According to the first manufacturer, the NPs were produced using an innovative patented method for physically obtaining non-ionic precious and semi-precious metal NPs. The process is based on a method of physically obtaining NPs through the laboratory disintegration of pure metals into particles a few to several nanometers in size.
A quality certificate for each batch of NPs (e.g., concentration in ppm, pH, shelf life, and batch number) was delivered by the manufacturer as confirmation that the NPs were of the necessary quality to conduct studies. The stability of the AgNPs, CuNPs, and AuNPs was also confirmed by the manufacturer.
2.2. Physicochemical Properties of Metal NPs
The zeta potential and the distribution of the AuNPs’ and NP-FeCs’ sizes were determined, as these NPs had not been included in previous research; a Zetasizer Nano ZS (ZEN3500; Malvern Instruments, Malvern, UK) was used for this purpose. The zeta potential, mobility, and conductivity of the AuNPs and NP-FeCs at concentrations of 50 mg/L were also obtained.
Dynamic laser scattering under electrophoretic conditions was used to measure the zeta potential, and the Smoluchowski correction was used to calculate the zeta potential. Each sample was measured after 120 s of stabilization at 25 °C in triplicate. Each replication contained at least 20 averaged analyses. Representative repetitions of the zeta potential distribution and mobility are presented in Figure 1A–D.
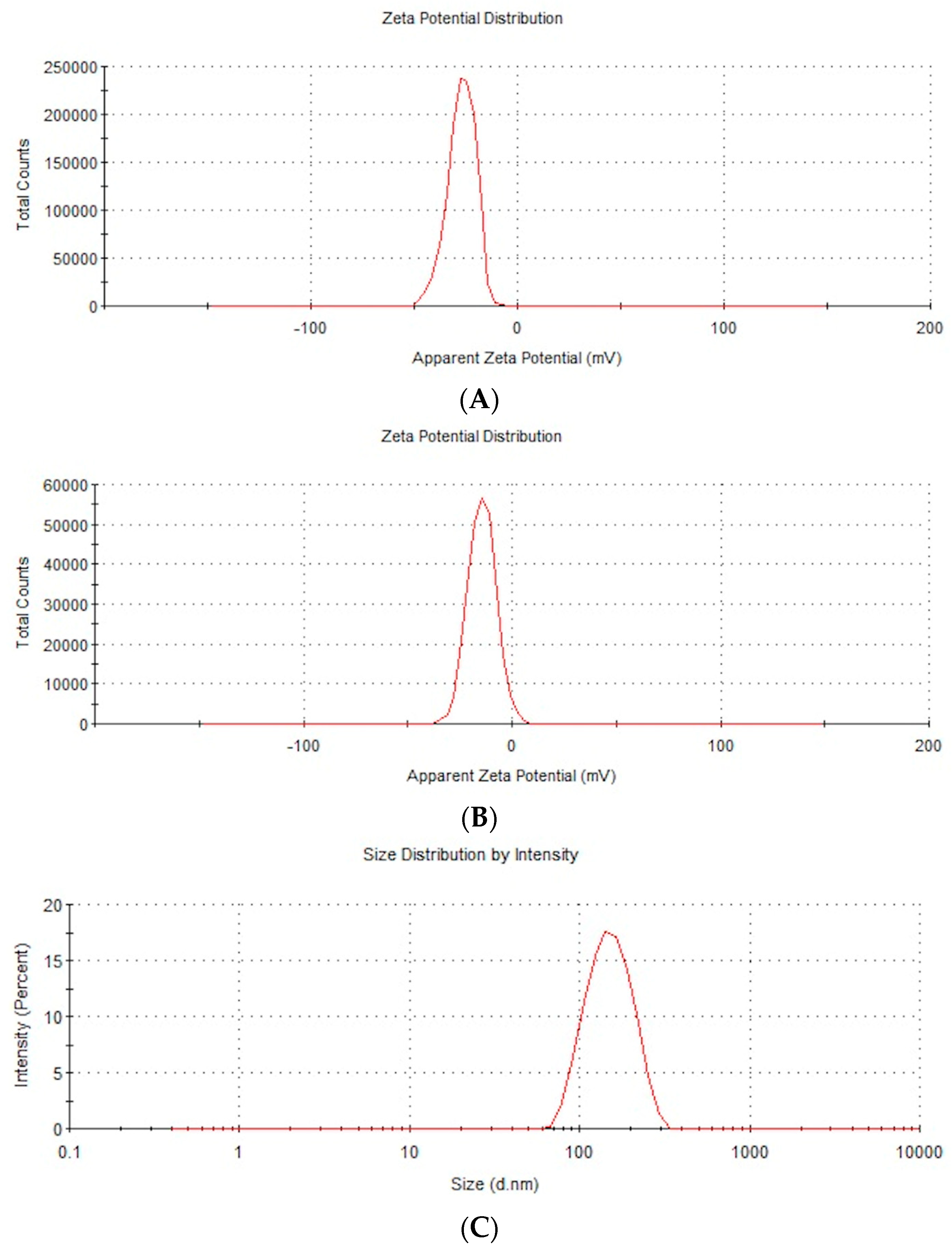
Figure 1. (A) Representative distribution of the AuNPs’ zeta potential at a concentration of 50 mg/L. (B) Representative distribution of the NP-FeCs’ zeta potential at a concentration of 50 mg/L. (C) Representative distribution of the AuNPs’ size at a concentration of 50 mg/L. (D) Representative distribution of the NP-FeCs’ size at a concentration of 50 mg/L.
The size and structure distribution of the nanoparticles was determined by analyzing transmission electron microscopy images and hydrodynamic size measurements. The NPs were observed using a JEM-2000EX (JEOL, Tokyo, Japan) transmission electron microscope. The NPs, in hydrocolloid form, were sonicated for 60 min, then applied to the forming mesh, and left to dry for 24 h. Images of the AuNPs and NP-FeCs were taken at a voltage of 200 kV. In order to analyze the hydrodynamic size of the NPs, they were sonicated for 30 min and then measured using a Zetasizer Nano ZS (ZEN3500; Malvern Instruments, Malvern, UK). The hydrodynamic size was determined using dynamic laser light scattering (DLS). Each sample was measured after 120 s of stabilization at 25 °C in triplicate. Each replication contained at least 10 averaged analyses.
2.3. The In Vitro Culture of BME-UV1 Cells
Bovine mammary gland cells of the BME-UV1 line used in the experiment were provided by M. Gajewska (Veterinary Medicine Faculty, Warsaw University of Life Sciences). The BME-UV1 cells were kept in appropriate DMEM/F12 medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% bovine serum and with antibiotic and antimycotic agents added (Gibco™ Antibiotic-Antimycotic, Thermo Fisher Scientific, Waltham, MA, USA). The cells were cultured in vitro in bottles for adherent cell culture at a temperature of 37 °C in an atmosphere with 5% CO2 in a NuAire DH AutoFlow CO2 incubator (Plymouth, MN, USA).
2.4. The Viability of BME-UV1 Cells after Incubation with AuNPs and NP-FeCs
AuNP and NP-FeC solutions were prepared according to the methods described in this paper in Section 2.1.
The viability of the BME-UV1 cells was calculated using the PrestoBlue test (Thermo Fisher Scientific, USA). The PrestoBlue test’s reagent is a ready-to-use resazurin-based solution for which cells are permeable. It can, therefore, be useful as a cell viability indicator using the reducing power of living cells. The reagent is modified by reducing the environment of viable cells and turns from blue to purple or pink in color, becoming highly fluorescent. Due to this phenomenon, the color changes can be evaluated using absorbance or fluorescence values.
BME-UV1 cells were placed in sterile 96-well plates with covers at 5 × 103 cells per well. The cells were incubated for 24 h. Next, the medium was removed and 90 μL of AuNP and NP-FeC hydrocolloids at a concentration of 0.5, 1, 2.5, or 5 mg/L were added to each well. The control group consisted of cells kept in the medium without the addition of NPs. After 24 h of incubation, 10 μL of PrestoBlue reagent was added to each well and the prepared plates were incubated for 2 h at 37 °C. The absorbance value was measured at a wavelength of 570 nm in an Infinite M200 immunoenzymatic reader (Tecan, Durham, NC, USA).
2.5. The In Vitro Membrane Integrity of BME-UV1 Cells after Incubation with AuNPs and NP-FeCs
The amount of lactate dehydrogenase (LDH) was verified in order to evaluate the membrane integrity of the BME-UV1 cells. The assay can be used to quantitatively measure the extracellular LDH release in culture media, due to a specific enzymatic reaction. The result of the process is a red formazan product that can be spectrophotometrically measured.
Cells of the BME-UV1 line were placed in 96-well plates at 5 × 103 cells per well. The plates were incubated for 24 h. Following this, the medium was removed and 90 μL of AuNP and NP-FeC hydrocolloids at a concentration of 0.5, 1, 2.5, or 5 mg/L were added to each well. The control group consisted of cells placed in the medium without the addition of NPs. After 24 h of incubation, the plates were centrifuged and 50 μL of the medium was placed in new 96-well plates. An LDH working solution was then added to each well according to the producer’s instructions (LDH Cytotoxicity Assay Kit, Thermo Fisher Scientific). The plates were incubated for 20 min at room temperature without light. In the next step, the reaction was completed by adding 50 μL of a stop solution. Absorbance values were measured at a wavelength of 490 nm in an Infinite M200 immunoenzymatic reader (Tecan, Durham, NC, USA).
2.6. The In Vitro Estimation of the Number of BME-UV1 Cells after Incubation with AuNPs and NP-FeCs
In order to assess the number of BME-UV1 cells, the amount of lactate dehydrogenase (LDH) released from all cells after they were lysed was evaluated. BME-UV1 cells were placed in 96-well plates at 5 × 103 cells per well. The plates were incubated for 24 h. After this, the medium was removed and 90 μL of AuNP and NP-FeC hydrocolloids at a concentration of 0.5, 1, 2.5, or 5 mg/L were added per well. The control group consisted of cells placed in the medium without the addition of NPs. After 24 h of incubation, the plates were centrifuged and 50 μL of the medium was placed in new 96-well plates. An LDH working solution was then added to each well according to the producer’s instructions (LDH Cytotoxicity Assay Kit, Thermo Fisher Scientific). The plates were incubated for 20 min at room temperature without light. In the next step, the reaction was completed by adding 50 μL of a stop solution. Absorbance values were measured at a wavelength of 490 nm in an Infinite M200 immunoenzymatic reader (Tecan, Durham, NC, USA) and normalized to a measurement of 680 nm.
2.7. Statistical Analysis
The obtained results were analyzed using one-way analysis of variance (ANOVA) in the Prism 8 program (GraphPad Software v. 4.0, La Jolla, CA, USA). Differences between groups were estimated using Tukey’s test. The results of the survey were presented as average values with standard deviations. Differences at p ≤ 0.05 were considered to be statistically significant.
2.8. In Vitro Bacterial and Fungi Cultures
Several species of bacteria from different groups were obtained from LGC Standards (Łomianki, Poland). The examined pathogens were as follows: Gram-positive bacteria (Staphylococcus aureus, Streptococcus agalactiae, Streptococcus uberis), Gram-negative bacteria (Enterococcus faecalis, Escherichia coli, Enterobacter cloacae, Pseudomonas aeruginosa), and the yeast Candida albicans. The cultured microorganisms were kept in 20% glycerol solution under temperature conditions of −20 °C.
The pathogens were thawed and rinsed with sterile distilled water in order to remove the glycerol. In the next step, bacterial cells or fungi were added to nutrient broth medium (Bio-Rad, Warsaw, Poland) that had been sterilized in glass flasks in a Classic 2100 autoclave (Prestige Medical, Chesterfield, UK). The flasks were placed in a rotating incubator at a temperature of 37 °C (Stuart SI500 Vernon Hills, IL, USA).
2.9. Preliminary NP Concentrations
Bacterial cells pipetted from the cell culture and incubated for one night (at 37 °C) were used in the experiments. The experimental groups contained nutrient broth and AgNPs, CuNPs, or AgCuNPs at a concentration of 0.5, 1, or 2.5 µg/mL. The control group contained nutrient broth without the addition of NPs. Each group was prepared in three repetitions. In the next step, 100 μL of the microorganism species (at a concentration of 1 × 106 microorganisms per mL) mentioned earlier were added to prepared flasks. The samples were incubated for 24 h in a rotating incubator (SI500) at a temperature of 37 °C and a rotation speed of 70 revolutions per minute. The microorganisms’ viability was calculated using the PrestoBlue test. After incubation, 90 µL of the medium was placed in 96-well plates and 10 µL of PrestoBlue reagent (Thermo Fisher Scientific) was added to each well. Each sample was placed in the well in six repetitions. The plates were incubated for 20 min at 37 °C. Absorbance was measured at a wavelength of 570 nm in an Infinite M200 immunoenzymatic reader (Tecan). The viability of the pathogens was presented as a percentage of the viability of the control group, according to the following equation:
2.10. Preparation of Mixture and Wax with the Addition of NPs
The fluid mixture consisted of 10% vegetable glycerin and 10% propylene glycol in distilled water with the addition of NPs at selected concentrations. The mixture included homogenous NPs or an AgCuAuNP complex prepared according to the description presented in Section 2.1. The NP hydrocolloids consisting of homogenous NPs or the AgCuAuNP complex were mixed with vegetable glycerin and propylene glycol in appropriate proportions at 25 °C and vortexed for 60 s.
The beeswax and olive oil mixture was heated at 37 °C to a uniform consistency. The NP hydrocolloids consisting of homogenous NPs or the AgCuAuNP complex were mixed with the prepared wax and vortexed for 5 min at 37 °C.
3. Results
The size distribution was determined by analyzing images of AuNPs and NP-FeCs obtained using a transmission electron microscope, as was the hydrodynamic size (average size of agglomerates). Measurements of the hydrodynamic size were carried out using hydrocolloids at a concentration of 50 mg/L. The results of the hydrodynamic size measurements and the sizes of the nanoparticles measured using transmission electron microscopy images are shown in . In addition, the structure of the AuNPs and NP-FeCs was determined based on images obtained using electron microscopy. Selected microphotographs are shown in Figure 2 and Figure 3.

Figure 2. The microphotographs of the AuNPs obtained using a transmission electron microscope.
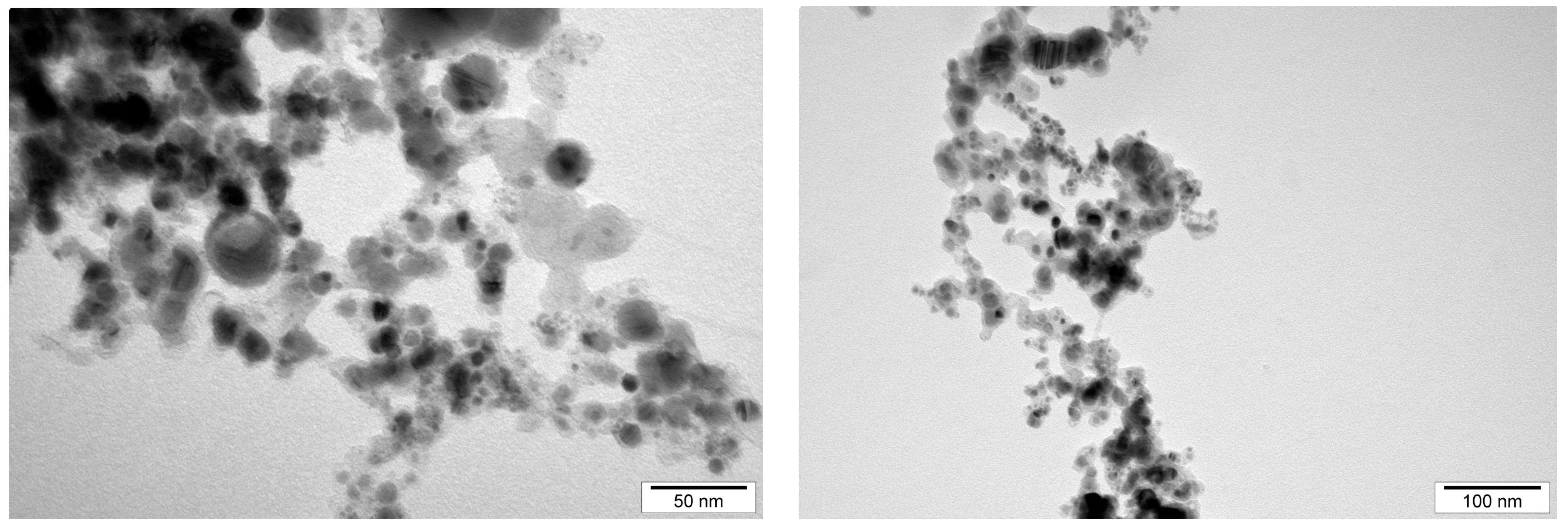
Figure 3. The microphotographs of the NP-FeCs obtained using a transmission electron microscope.
3.1. The Estimated Viability of the BME-UV1 Cells after Incubation with AuNPs and NP-FeCs
The in vitro viability of the BME-UV1 cells was determined using a PrestoBlue assay, measuring the redox activity of the cells. AuNPs at concentrations of 0.5, 1, 2.5, and 5 mg/L had no cytotoxic effect on the epithelial mammary cells (BME-UV1). The viability of the selected cell line decreased only when cells were treated with NP-FeCs at the highest concentration (5 mg/L). The results for cell viability after 24 h of incubation with AuNP and NP-FeC hydrocolloids are shown in Figure 4A. NP-FeCs at concentrations of 0.5, 1, and 2.5 mg/L had no cytotoxic effect on the epithelial mammary cells (BME-UV1).
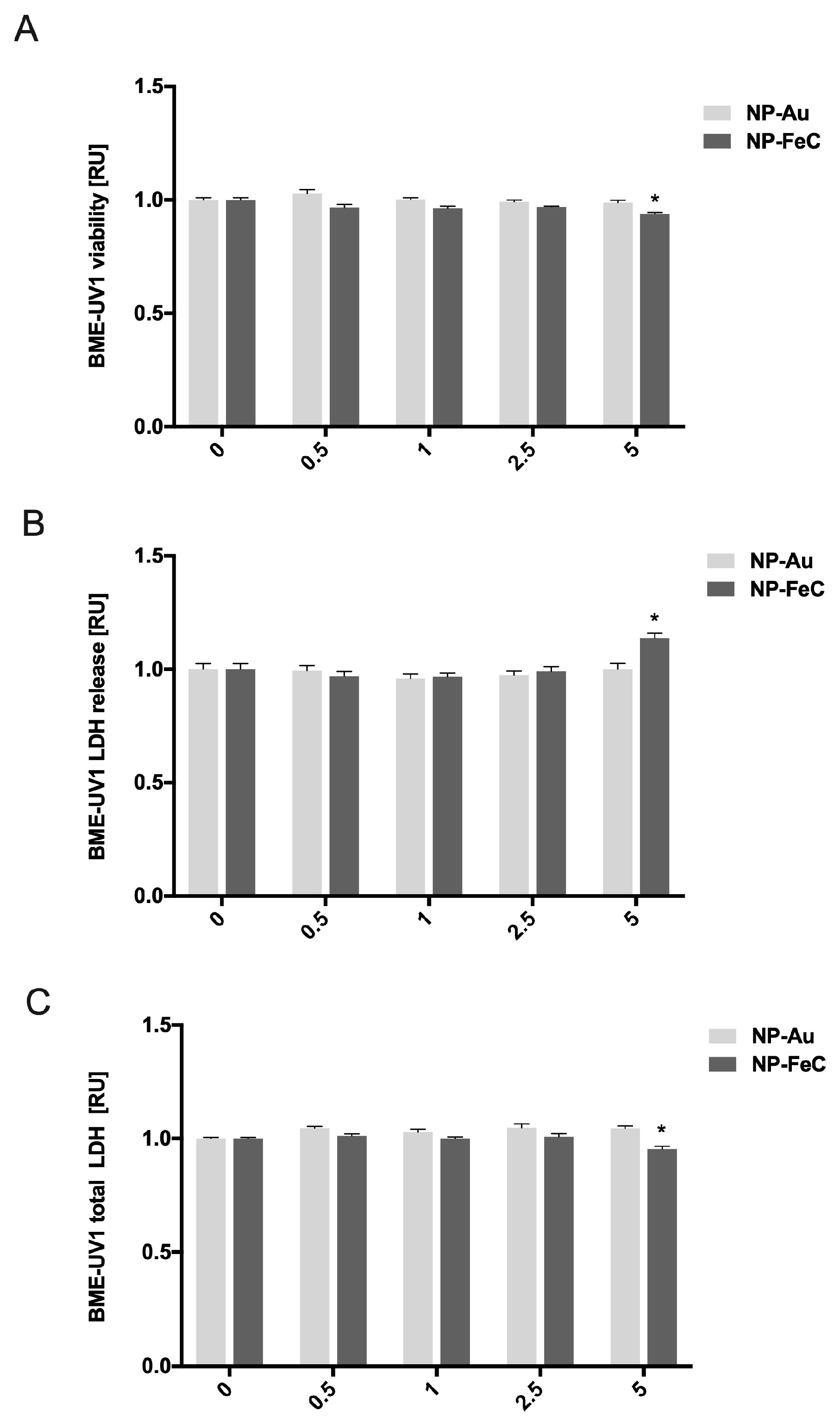
Figure 4. The viability of BME-UV1 cells estimated using a PrestoBlue assay (A). Evaluation of BME-UV1 cell membrane integrity (B). Determination of the total number of BME-UV1 cells using the LDH assay (C). * Statistically significant differences at p < 0.05.
The integrity of the BME-UV1 cells’ membranes was assessed using the LDH Cytotoxicity Assay Kit (Thermo Fisher Scientific, USA). The cell membrane’s integrity was compromised after treatment with NP-FeCs at a concentration of 5 mg/L. The results for the cell membrane’s integrity after 24 h of incubation with AuNP and NP-FeC hydrocolloids are shown in Figure 4B.
The total number of BME-UV1 cells was assessed using the LDH Cytotoxicity Assay Kit (Thermo Fisher Scientific, USA) by determining the total amount of LDH. The number of cells changed after being treated with NP-FeCs at a concentration of 5 mg/L. The results for the cell counts after 24 h of incubation with AuNP and NP-FeC hydrocolloids are shown in Figure 4C.
3.2. The Cytotoxic Effect of AgNPs, CuNPs, AuNPs, and NP-FeCs on Pathogen Viability
The obtained results suggested that metal NPs can decrease the viability of the selected mastitis bacteria, Staphylococcus aureus, Streptococcus agalactiae, Streptococcus uberis, Enterococcus faecalis, Escherichia coli, Enterobacter cloacae, and Pseudomonas aeruginosa, and the yeast Candida albicans even at low concentrations . The results included the average viability for each group and the standard error (SE) after incubation with homogenous NP hydrocolloids at various concentrations (0.5–5 ppm). The presented data were statistically significant for all the examined groups (p < 0.01).
The viability of the pathogens used in the experiment decreased in three of the experimental groups: AgNPs, CuNPs, and AuNPs.
The strongest antimicrobial properties—decreasing the viability of S. uberis, S. agalactiae, and E. faecalis by 50% or more—were observed for AgNPs at a concentration of 5 ppm. The results for E. cloacae pointed to the strongest biocidal effect having occurred in the AuNPs5 group (AuNPs at a concentration of 5 ppm; see . Similar results were observed in the CuNPs5 group for S. uberis and E. faecalis, where the viability of the pathogens decreased by 45%.
Conversely, the group with NP-FeCs added promoted bacterial growth, which was indicated by viability values >100% (the control group’s viability was always 100%). The exception was the S. aureus group, where the viability varied from 96.05% to 99.97%. Therefore, NP-FeCs were not included in later parts of the experiment because of their lack of antimicrobial properties.
3.3. The Cytotoxic Effect of the AgCuAuNP Complex on Pathogen Viability
The cytotoxic effect of the AgCuAuNP complex was observed during the experiment . The combination of the three different metal NP types produced lower viabilities at all concentrations for all tested pathogens. The NPs’ strongest biocidal effect was observed at the highest concentration (5 ppm). The difference between the 2 ppm and 5 ppm concentrations was relatively small in the case of the selected pathogen species. The presented data were statistically significant for all the examined groups (p < 0.01).
3.4. The In Vitro Antimicrobial Properties and Cytotoxic Effect of the AgCuAuNP Complex
The wax that included homogenous NPs had lower antimicrobial properties, decreasing the pathogens’ viability by 14–20% . The exception was the wax with homogenous NPs applied to S. agalactiae, which decreased the viability by 47% in the case of AgNPs and AuNPs. The AgCuAuNP complex had the weakest antibacterial activity against S. aureus (71%). The viability of E. coli, S. agalactiae, and C. albicans was 55%, 57%, and 66%, respectively.
The results of adding homogenous NPs to the liquid mixture were similar to those for the wax groups. Similar data were obtained for the AgCuAuNP complex, with the strongest antimicrobial activity being observed in the E. coli group. The obtained results were statistically significant for all groups (p < 0.01).
4. Discussion
4.1. Nanotechnology and NPs’ Properties
Inflammation of the tissue of the bovine mammary gland is still the most common disease found in high-yielding dairy cattle. It causes significant deterioration of the cows’ welfare and affects the parameters and cytological quality of milk [11].
The antimicrobial properties of metal NPs are associated with various factors, including their stability in hydrocolloid form, which results in no agglomeration phenomenon and stronger biocidal properties because the NPs’ surfaces are not limited [12]. In this study, AuNPs and NP-FeCs had zeta potential values of −28.4 mV and −18.5 mV, respectively. These results suggest weak antibacterial properties, because the limit of NP stability is ±30 mV. Hence, the average size of the agglomerates was higher for NP-FeCs than for AuNPs (342.9 nm vs. 148.3 nm, respectively). The physicochemical properties of AgNPs, AuNPs, and CuNPs, obtained from commercial sources have been confirmed in previous studies [13,14,15].
4.2. The Influence of NPs on the Viability of Bovine Mammary Gland Cells
The toxic effect of NPs on water organisms has been observed in several papers [16,17,18]. The exposure of water organisms to NPs usually causes several problems, for example, effects on larval development, impaired reproduction, or increased oxidative stress [17,18]. Moreover, for aquatic animals, AgNPs are often considered to be more toxic than CuNPs [16]. At the same time, NPs are used in veterinary and human medicine, for example, in cancer treatment or disinfectant products. An interesting phenomenon was described by Matuszewski et al. [19], proving that in ovo inoculation with calcium carbonate NPs positively influences bone mineralization if the process takes place during chicken embryogenesis [19]. The described correlations have not been completely examined, and further studies—both in vitro and in vivo—are necessary to evaluate the long-term effects of NPs on animals and humans.
Studies focusing on the long-term effects of NPs on bovine cells are surprisingly limited [13,20]. The safety of animals and humans is a crucial issue if NPs are to be used as an alternative antimicrobial agent in the future. According to Jagielski et al. [20], AgNPs have no cytotoxic effect on mammary gland cells if used at concentrations of up to 1–4 mg/L [20]. Similar results were obtained by Kalińska et al. [13]. However, study results vary among scientists and some papers point out that NPs can be toxic for humans [21]. Nevertheless, available references are still limited in the case of the BME-UV1 cell line. Kalińska et al. [13] revealed that AgNP, AgCuNP, and CuNP concentrations of 0.5–2.5 mg/L increase the likelihood of spontaneous leaking of LDH by 20–60% (p < 0.05). The authors of this paper did not observe an increase in LDH release in the case of AuNPs and NP-FeCs, and no negative impact on BME-UV1 cells was observed. The use of an LDH assay can be a valuable factor in assessing the potential toxicity of new antimicrobial agents, as cellular proteins can adhere to NPs as a protein corona. But the LDH assay should always be carefully included for evaluating the NPs’ effect on cell viability [22].
4.3. The Antimicrobial Properties of NPs
The increasing number of strains that are resistant to antibiotics is a clearly observed phenomenon [7,23]. Scientists are therefore struggling to invent new agents that could be used as alternatives to drug treatments [2]. Another important issue in current studies is the development of rapid diagnostic methods for mastitis that are accurate regarding at least the type of pathogen and preferably the species, for example, Staphylococcus aureus. This would contribute significantly to speeding up the time for diagnosis and the implementation of appropriate treatment. For this reason, markers for udder inflammation in cows are still being investigated. Some studies have indicated that whey proteins may act as markers [24], but there are no clear bovine markers that are currently considered to be indicators of developing inflammation in the udder. Somatic cell count (SCC) growth is still the most commonly used symptom for the diagnosis of subclinical inflammation. Some of the most promising solutions for prophylaxis of inflammation are plant extracts, fungi extracts, phage therapy, lysozyme, lactoferrin, bacteriocins, and NPs [13,24,25,26,27,28].
Previous studies have revealed that NPs can be used in cases of several bovine diseases, for example, mastitis [13] and lameness [14]. This solution is also promising due to the fact that NPs do not negatively influence bovine mammary tissue [14]. Another important issue is whether it is possible to obtain high-quality NPs from commercial sources [13,14], which is an important factor in planning the further development of NP products. Studies using NPs in pre-milking and post-milking disinfectant products are limited, but in vitro experiments suggest that AgNPs and CuNPs can reduce pathogen viability by 50% [16]. The antimicrobial properties of some metal NPs are well known, for example, AgNPs and CuNPs [13,14,15,26]; the idea of using the synergistic effect of at least two different NPs has also previously been presented [13,26]. In the case of mastitis, AuNPs, PtNPs, and NP-FeCs demonstrate weaker antimicrobial properties even at high concentrations [14,29], and these results are similar to the data presented in this paper.
4.4. Teat Sealant in Dry Cow Therapy
The authors of this study estimate the average cost of mastitis incidences to be a minimum of €240/cow/year [30]. Similar data from other European countries estimate that the total mastitis cost per cow per year ranges from €261 to €483 [31,32]. In contrast, estimated average losses due to mastitis in the U.S. are at a minimal level of $131/cow/year [33].
Dry cow therapy is a crucial point in the production cycle. In accordance with EU Regulation 2019/6 on veterinary medicinal products that repealed Directive 2001/82/EC of 28 January 2022, regulations force veterinarians to reduce the use of antibiotics in animal production and are banning the use of some antibiotics as a method of prophylaxis. The amount of antibiotics used in the EU is highly disturbing, with an average of 84.4 mg/PCU (standardization unit), with some countries having almost double this value, for example, Cyprus, Poland, Hungary, Italy, Spain, and Portugal [34]. In addition, the guidelines in the Green Deal Industrial Plan indicate the need to reduce antibiotic use by 50% as early as 2030 [34,35].
In conclusion, the use of antibiotics as the most common form of mastitis treatment in dairy cows also requires a change in approach to reduce antibiotic use. One of the most popular procedures is combining antibiotic treatment with teat sealants during dry cow therapy. The first teat sealants were created in the early 1970s and were based on bismuth subnitrate; it is still the most common substance used. There are several papers proving that the use of teat sealants on dry cows, in addition to antibiotic treatment, has a positive influence, resulting in lower SCCs at the beginning of lactation and even higher milk production [36,37,38,39].
Based on the results obtained from the in vitro testing of wax plugs and teat disinfectant liquid, a decrease can be observed in the viability of the pathogens that most often cause mastitis in cows, resulting in a decrease in SCCs in milk and an increase in the milk yield of cows. ElAshmawy et al. [40] showed that there was an increase in milk production (by 1.84 kg/day) in cows treated with antibiotics and intramammary sealants compared to the control group. The authors also demonstrated a reduction in the SCCs during the first 150 days of lactation in cows treated with antibiotics and sealants and with sealants alone compared to controls. Molina et al. [37], in their study, used a combination of the antibiotic cloxacillin and an injectable sealant containing bismuth nitrite in its formulation. The authors showed that there was a decrease in the incidence of new mastitis infections by 8% in the early postpartum period after using the antibiotic and sealant compared to controls using the antibiotic alone. It was also observed that there was a reduction in the amount of E. coli bacteria in the milk of cows after calving compared to the pre-delivery sample after using the sealant. In contrast, the use of injectable sealants had no effect on the frequency of streptococcal isolation. Clabby et al. [41] carried out an experiment in which they dried one experimental group of cows with dystrophin sealants and an antibiotic and the other with the dystrophin sealant alone. The authors found that cows dried with the teat sealant alone had higher SCCs and were more likely to develop udder infections during the next lactation compared to cows dried with both the antibiotic and the sealant.
In conclusion, it is likely that NPs have high potential to be biocidal agents in future mastitis treatment and prevention also in dried cows. However, the obtained in vitro results require further studies using strains isolated from cows with clinical mastitis. Thus, the next step should be experiments with pathogens isolated from cows with mastitis, since these strains (including S. aureus or E. coli) may have different resistance to the tested NPs. For this reason, it is necessary to verify their biocidal properties also against field strains.
5. Conclusions
Limiting the phenomenon of antibiotic resistance is mainly related to discontinuing the use of antibiotics as a prophylaxis method. Before beginning treatment, it is necessary to identify the etiological factor so the appropriate drug can be selected in the appropriate dosage. Sales of antibiotics in some countries continue to increase, especially in the livestock sector, which poses a serious threat to the health of consumers. In order to prevent the increasing problem of antibiotic resistance in animal livestock, it is necessary to look for alternative agents and forms of treatment that will significantly reduce the use of antibiotics in the future. The conducted in vitro studies indicate that the use of AuNPs, AgNPs, CuNPs, and their triple complexes reduces the viability of the most common mastitis pathogens, i.e., S. aureus, E. coli, and S. uberis. Moreover, these NPs are not toxic to bovine mammary gland tissue. This suggests that in the future, NPs could be a new alternative in the prevention and treatment of mastitis in dry cows, but further in vivo studies are needed.
References
- Ruegg, P.L. A 100-Year Review: Mastitis detection, management, and prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [PubMed]
- Kalińska, A.; Gołębiewski, M.; Wójcik, A. Mastitis pathogens in dairy cattle—A review. World Sci. News 2017, 89, 22–31. [Google Scholar]
- Cobirka, M.; Tancin, V.; Slama, P. Epidemiology and Classification of Mastitis. Animals 2020, 10, 2212. [Google Scholar] [CrossRef]
- Sukumar, K.; James, P.C. Incidence of fungal mastitis in cattle. Tamilnadu J. Vet. Anim. Sci. 2012, 8, 356–359. [Google Scholar]
- Schwarz, S.; Kehrenberg, C.; Walsh, T.R. Use of antimicrobial agents in veterinary medicine and food animal production. Int. J. Antimicrob. Agents 2001, 17, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Singer, R.S.; Finch, R.; Wegener, H.C.; Bywater, R.; Walters, J.; Lipsitch, M. Antibiotic resistance—The interplay between antibiotic use in animals and human beings. Lancet Infect. Dis. 2003, 3, 47–51. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 1–24. [Google Scholar] [CrossRef]
- Krömker, V.; Leimbach, S. Mastitis treatment—Reduction in antibiotic usage in dairy cows. Reprod. Domest. Anim. 2017, 52, 21–29. [Google Scholar] [CrossRef]
- Høiby, N.; Ciofu, O.; Johansen, H.K.; Song, Z.; Moser, C.; Jensen, P.Ø.; Molin, S.; Givskov, M.; Tolker-Nielsen, T.; Bjarnsholt, T. The clinical impact of bacterial biofilms. Int. J. Oral Sci. 2011, 3, 55–65. [Google Scholar] [CrossRef]
- Durán, N.; Marcato, P.D.; Conti, R.D.; Alves, O.L.; Costa, T.M.; Brocchi, M. Potential Use of Silver Nanoparticles on Pathogenic Bacteria, their Toxicity and Possible Mechanisms of Action. J. Braz. Chem. Soc. 2010, 21, 949–959. [Google Scholar] [CrossRef]
- Miglior, F.; Fleming, A.; Malchiodi, F.; Brito, L.F.; Martin, P.; Baes, C.F. A 100-Year Review: Identification and genetic selection of economically important traits in dairy cattle. J. Dairy Sci. 2017, 100, 10251–10271. [Google Scholar] [CrossRef] [PubMed]
- Khodashenas, B. The Influential Factors on Antibacterial Behaviour of Copper and Silver Nanoparticles. Indian Chem. Eng. 2016, 58, 224–239. [Google Scholar] [CrossRef]
- Kalińska, A.; Jaworski, S.; Wierzbicki, M.; Gołębiewski, M. Silver and Copper Nanoparticles—An Alternative in Future Mastitis Treatment and Prevention? Int. J. Mol. Sci. 2019, 20, 1672. [Google Scholar] [CrossRef] [PubMed]
- Kot, M.; Kalińska, A.; Jaworski, S.; Wierzbicki, M.; Smulski, S.; Gołębiewski, M. In Vitro Studies of Nanoparticles as a Potentially New Antimicrobial Agent for the Prevention and Treatment of Lameness and Digital Dermatitis in Cattle. Int. J. Mol. Sci. 2023, 24, 6146. [Google Scholar] [CrossRef] [PubMed]
- Kalińska, A.; Jaworski, S.; Wierzbicki, M.; Kot, M.; Radzikowski, D.; Smulski, S.; Gołębiewski, M. Silver and Copper Nanoparticles as the New Biocidal Agents Used in Pre- and Post-Milking Disinfectants with the Addition of Cosmetic Substrates in Dairy Cows. Int. J. Mol. Sci. 2023, 24, 1658. [Google Scholar] [CrossRef]
- Ostaszewska, T.; Chojnacki, M.; Kamaszewski, M.; Sawosz-Chwalibóg, E. Histopathological effects of silver and copper nanoparticles on the epidermis, gills, and liver of Siberian sturgeon. Environ. Sci. Pollut. Res. 2016, 23, 1621–1633. [Google Scholar] [CrossRef]
- Ostaszewska, T.; Śliwiński, J.; Kamaszewski, M.; Sysa, P.; Chojnacki, M. Cytotoxicity of silver and copper nanoparticles on rainbow trout (Oncorhynchus mykiss) hepatocytes. Environ. Sci. Pollut. Res. 2017, 25, 908–915. [Google Scholar] [CrossRef]
- Szudrowicz, H.; Kamaszewski, M.; Adamski, A.; Skrobisz, M.; Frankowska-Łukawska, J.; Wójcik, M.; Bochenek, J.; Kawalski, K.; Martynow, J.; Bujarski, P.; et al. The Effects of Seven-Day Exposure to Silver Nanoparticles on Fertility and Homeostasis of Zebrafish (Danio rerio). Int. J. Mol. Sci. 2022, 23, 11239. [Google Scholar] [CrossRef]
- Matuszewski, A.; Łukasiewicz, M.; Niemiec, J.; Kamaszewski, M.; Jaworski, S.; Domino, M.; Jasiński, T.; Chwalibóg, A.; Sawosz, E. Calcium Carbonate nanoparticles—Toxicity and effect of in ovo inoculation on chicken embryo development, broiler performance and bone status. Animals 2021, 11, 932. [Google Scholar] [CrossRef]
- Jagielski, T.; Bakuła, Z.; Pleń, M.; Kamiński, M.; Nowakowska, J.; Bielecki, J.; Wolska, K.; Grudniak, A.M. The activity of silver nanoparticles against microalgae of the Prototheca genus. Nanomedicine 2018, 13, 1025–1036. [Google Scholar] [CrossRef]
- Ahamed, M.; AlSalhi, M.S.; Siddiqui, M.K.J. Silver nanoparticle applications and human health. Clin. Chim. Acta 2010, 411, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-J.; Kim, H.; Liu, Y.; Han, H.-K.; Kwon, K.; Chang, K.-H.; Park, K.; Kim, Y.; Shim, K.; An, S.S.A.; et al. Incompatibility of silver nanoparticles with lactate dehydrogenase leakage assay for cellular viability test is attributed to protein binding and reactive oxygen species generation. Toxicol. Lett. 2014, 225, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Oliver, S.P.; Murinda, S.E. Antimicrobial resistance of mastitis pathogens. Vet. Clin. Food Anim. Pract. 2012, 28, 165–185. [Google Scholar] [CrossRef] [PubMed]
- Puppel, K.; Kalińska, A.; Kot, M.; Slósarz, J.; Kunowska-Slósarz, M.; Grodkowski, G.; Kuczyńska, B.; Solarczyk, P.; Przysucha, T.; Gołębiewski, M. The Effect of Staphylococcus spp., Streptococcus spp. and Enterobacteriaceae on the Development of Whey Protein Levels and Oxidative Stress Markers in Cows with Diagnosed Mastitis. Animals 2020, 10, 1591. [Google Scholar] [CrossRef]
- Radzikowski, D.; Kalińska, A.; Ostaszewska, U.; Gołębiewski, M. Alternative solutions to antibiotics in mastitis treatment for dairy cows—A review. Anim. Sci. Pap. Rep. 2020, 38, 117–133. [Google Scholar]
- Lange, A.; Grzenia, A.; Wierzbicki, M.; Strojny-Cieslak, B.; Kalińska, A.; Gołębiewski, M.; Radzikowski, D.; Sawosz, E.; Jaworski, S. Silver and Copper Nanoparticles Inhibit Biofilm Formation by Mastitis Pathogens. Animals 2021, 11, 1884. [Google Scholar] [CrossRef]
- Tamrakar, S.; Nishida, M.; Amen, Y.; Tran, H.B.; Suhara, H.; Fukami, K.; Parajuli, G.P.; Shimizu, K. Antibacterial activity of Nepalese wild mushrooms against Staphylococcus aureus and Propionibacterium acnes. J. Wood Sci. 2017, 63, 379–387. [Google Scholar] [CrossRef]
- Zhao, D.L.; Wang, D.; Tian, X.Y.; Cao, F.; Li, Y.Q.; Zhang, C.S. Anti-phytopathogenic and cytotoxic activities of crude extracts and secondary metabolites of marine-derived fungi. Mar. Drugs 2018, 16, 36. [Google Scholar] [CrossRef]
- Wernicki, A.; Puchalski, A.; Urban-Chmiel, R.; Dec, M.; Stegierska, D.; Dudzic, A.; Wojcik, A. Antimicrobial properties of gold, silver, copper and platinum nanoparticles against selected microorganisms isolated from cases of mastitis in cattle. Med. Weter 2014, 70, 564–567. [Google Scholar]
- Gołębiewski, M. Evaluation of changes in technical and production indicators on farms covered by the “Healthy Cow” program. In Proceedings of the Conference “Heathy Cow” Oral Presentation, Warsaw University of Life Sciences, Warsaw, Poland, 4 December 2013. [Google Scholar]
- van Soest, F.J.; Santman-Berends, I.M.; Lam, T.J.; Hogeveen, H. Failure and preventive costs of mastitis on Dutch dairy farms. J. Dairy Sci. 2016, 99, 8365–8374. [Google Scholar] [CrossRef]
- Doehring, C.; Sundrum, A. The informative value of an overview on antibiotic consumption, treatment efficacy and cost of clinical mastitis at farm level. Prev. Vet. Med. 2019, 165, 63–70. [Google Scholar] [CrossRef]
- Hogeveen, H.; Steeneveld, W.; Wolf, C.A. Production diseases reduce the efficiency of dairy production: A review of the results, methods, and approaches regarding the economics of mastitis. Annu. Rev. Resour. Econ. 2019, 11, 289–312. [Google Scholar] [CrossRef]
- Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2021, European Medicines Agency, 2022, Luxemburg. Available online: https://www.ema.europa.eu/en/documents/report/sales-veterinary-antimicrobial-agents-31-european-countries-2021-trends-2010-2021-twelfth-esvac_en.pdf (accessed on 15 May 2023).
- Wiśniewski, P.; Marks-Bielska, R. The importance of the implementation of the European Green Deal for the Polish countryside and agriculture. In Polska Wieś 2022: Raport o Stanie Wsi; Jerzy, W., Andrzej, H., Eds.; Fundacja na rzecz Rozwoju Polskiego Rolnictwa: Warsaw, Poland, 2022; pp. 119–132. ISBN 978-83-66849-54-9. [Google Scholar]
- Meaney, W.J. Effect of a dry period teat seal on bovine udder infection. Ir. J. Agric. Res. 1977, 16, 293–299. [Google Scholar]
- Molina, L.R.; Costa, H.; Leão, J.M.; Malacco, V.M.; Facury, E.J.; Carvalho, A.U.; Lage, C.F. Efficacy of an internal teat seal associated with a dry cow intramammary antibiotic for prevention of intramammary infections in dairy cows during the dry and early lactation periods. Pesq. Vet. Bras. 2017, 37, 465–470. [Google Scholar] [CrossRef]
- Freu, G.; Tomazi, T.; Monteiro, C.P.; Barcelos, M.M.; Alves, B.G.; dos Santos, M.V. Internal teat sealant administered at drying off reduces intramammary infections during the dry and early lactation periods of dairy cows. Animals 2020, 10, 1522. [Google Scholar] [CrossRef]
- Lavery, A.; Craig, A.L.; Gordon, A.W.; Ferris, C.P. Impact of adopting non-antibiotic dry-cow therapy on the performance and udder health of dairy cows. Vet. Rec. 2022, 190, e1731. [Google Scholar] [CrossRef]
- ElAshmawy, W.R.; Okello, E.; Williams, D.R.; Anderson, R.J.; Karle, B.; Lehenbauer, T.W.; Aly, S.S. Effectiveness of Intramammary Antibiotics, Internal Teat Sealants, or Both at Dry-Off in Dairy Cows: Milk Production and Somatic Cell Count Outcomes. Vet. Sci. 2022, 9, 559. [Google Scholar] [CrossRef]
- Clabby, C.; McParland, S.; Dillon, P.; Arkins, S.; Flynn, J.; Murphy, J.; Boloña, P.S. Internal teat sealants alone or in combination with antibiotics at dry-off–the effect on udder health in dairy cows in five commercial herds. Animal 2022, 16, 100449. [Google Scholar] [CrossRef]